Cellular Energy: From Fuel to ATP
How a cell takes a molecule of sugar or fat from your bloodstream and turns it into usable energy — every transporter, every enzyme, every cofactor, in the order it actually happens.
Why this page exists
Almost every other topic on this site connects back to one question: is the cell making enough energy, and what is getting in the way? Fatigue, brain fog, slow metabolism, poor recovery, cold hands — these are very often downstream of a problem somewhere in the chain described below.
This page builds that chain from the ground up. We follow a single molecule of glucose from the blood, into the cell, into the mitochondria, and all the way to the finished product — and then we do the same for fat. By the end you should be able to picture the whole system like an engineer looking at a power plant: where the fuel enters, where it gets processed, where the bottlenecks are, and what each cofactor is doing.
Two definitions before we start, because everything depends on them:
-
ATP (adenosine triphosphate) is the cell's energy currency. It is a small molecule with three phosphate groups attached in a row. The bond holding the last phosphate on is energy-rich: when the cell snaps that phosphate off (turning ATP into ADP, adenosine diphosphate, with only two phosphates), energy is released that powers some piece of cellular work. The cell then re-attaches a phosphate to ADP to rebuild ATP, and the cycle repeats. You are not "burning" ATP the way you burn petrol — you are charging and discharging it like a rechargeable battery, millions of times per second.
-
A mitochondrion (plural: mitochondria) is a compartment inside the cell — its own little sealed bag with a double membrane — where the bulk of ATP is produced. Think of it as the cell's power station. A single cell can contain anywhere from a handful to several thousand mitochondria, depending on how energy-hungry it is. Heart muscle cells are packed with them; they make up about a third of the cell's volume.
Everything below is, ultimately, the story of how fuel becomes ATP inside or around these mitochondria.
The big picture, in one paragraph
Glucose enters the cell and is partially broken down in the main body of the cell (the cytosol) in a process called glycolysis, producing a little ATP and a molecule called pyruvate. Pyruvate is shipped into the mitochondria and converted into acetyl-CoA, which feeds into a loop of chemical reactions called the Krebs cycle. The Krebs cycle does not make much ATP directly. Instead, its real job is to strip high-energy electrons off the fuel and load them onto two carrier molecules, NADH and FADH₂. These carriers deliver their electrons to the electron transport chain — a series of protein complexes embedded in the inner mitochondrial membrane. As electrons pass down this chain, the energy released is used to pump protons (hydrogen ions) across the membrane, building up a kind of pressure. That pressure then drives a molecular turbine called ATP synthase, which spins and manufactures the bulk of the cell's ATP. Oxygen sits at the very end to collect the spent electrons. Fat enters the same system slightly differently but converges at acetyl-CoA.
That is the entire process. The rest of this page is that paragraph, slowly, with every part named and explained. Here it is as a map — refer back to it as we go:
flowchart TD
G[Glucose in blood] -->|GLUT doors| GC[Glucose in cytosol]
GC -->|Glycolysis<br/>net +2 ATP, +2 NADH| PYR[Pyruvate]
PYR -->|into mitochondria| PDH["PDH gate<br/>(needs vitamin B1)"]
PDH --> ACA[Acetyl-CoA]
FAT[Fatty acids in blood] -->|CD36 door| FC[Fatty acid in cytosol]
FC -->|"Carnitine shuttle<br/>(CPT1 gate)"| BOX[Beta-oxidation]
BOX --> ACA
ACA --> KREBS["Krebs cycle<br/>charges up NADH + FADH2"]
KREBS --> ETC["Electron transport chain<br/>pumps protons · uses oxygen"]
ETC --> ATPS[ATP synthase turbine]
ATPS --> ATP["ATP<br/>(~90% of the total)"]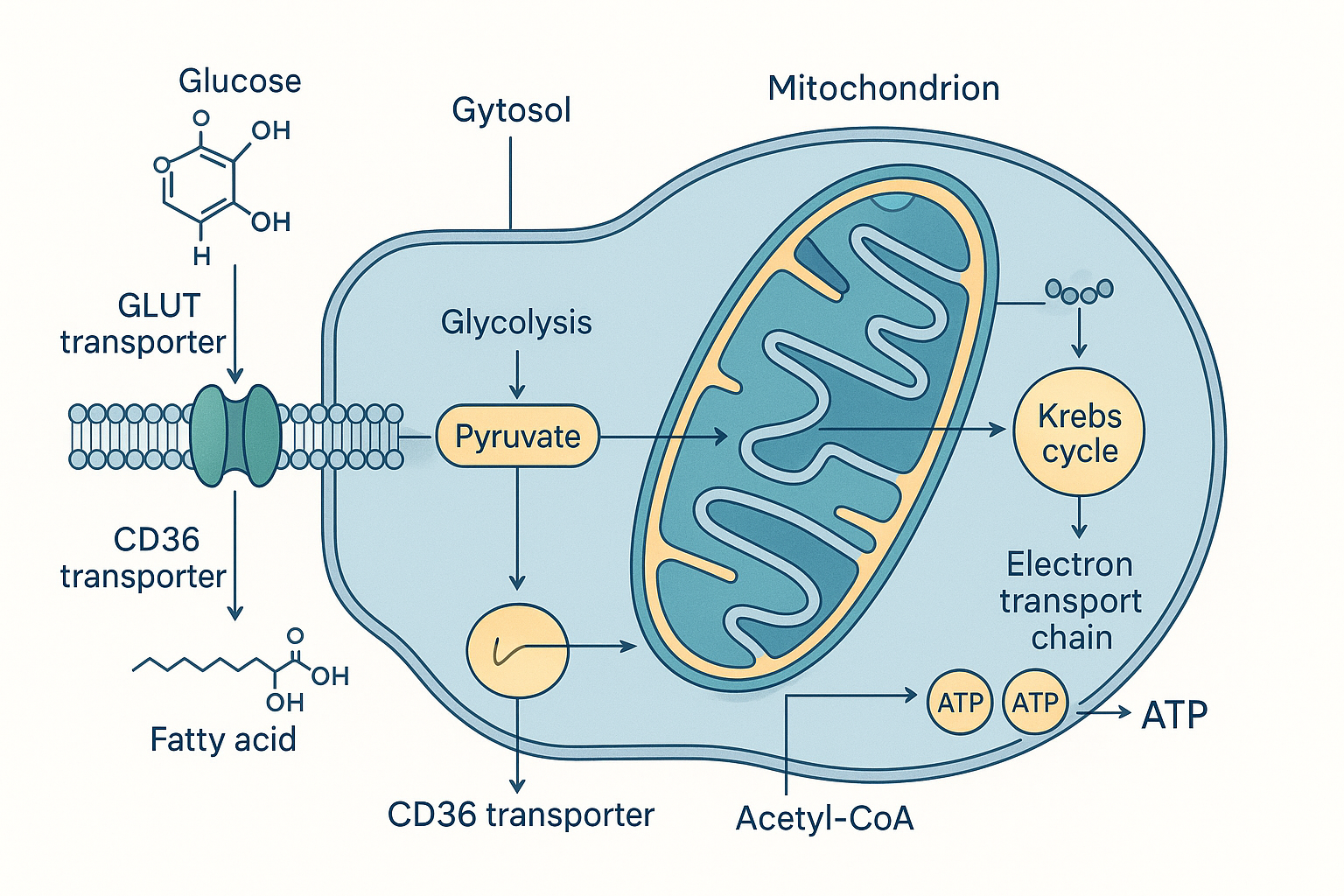 The whole journey in one picture: fuel enters the cell, the front-ends differ, both converge inside the mitochondrion, and ATP comes out.
The whole journey in one picture: fuel enters the cell, the front-ends differ, both converge inside the mitochondrion, and ATP comes out.
Part 1 — Glucose as fuel
Step 1: Getting glucose out of the blood and into the cell
Glucose is a sugar molecule dissolved in your blood. The inside of a cell is separated from the blood by the cell membrane — a thin double layer of fat (lipid) molecules. Glucose cannot simply drift through this fatty barrier, because glucose is water-loving (hydrophilic) and the membrane interior is fat-loving (hydrophobic). It needs a doorway.
Those doorways are proteins called GLUT transporters (GLUT = glucose transporter). They sit in the membrane and form a channel that glucose can pass through. There are several types, numbered GLUT1 through GLUT4, and the difference between them matters:
-
GLUT1 and GLUT3 are "always-on." They sit permanently in the membrane and let glucose trickle in continuously. The brain relies heavily on these, which is one reason the brain gets first claim on blood glucose regardless of hormonal state — it does not have to wait for permission.
-
GLUT2 is the low-affinity transporter of the liver, the gut lining, and the pancreas. It only moves glucose in bulk when blood sugar is high, which makes it a sensor and a buffer rather than a steady feeder. The liver uses it to soak up surplus glucose after a meal — and to let glucose back out when the body needs it — and the pancreas uses it to detect high blood sugar and release insulin. Unlike GLUT4, it needs no insulin to open.
-
GLUT4 is the regulated one, found mainly in muscle and fat tissue. GLUT4 normally sits hidden inside the cell, stored in little membrane bubbles. It only moves to the surface when instructed by the hormone insulin. When you eat carbohydrate, blood glucose rises, the pancreas releases insulin, insulin binds receptors on muscle and fat cells, and those cells respond by shuttling GLUT4 to their surface — opening the doors so glucose can flood in. This is the central event that insulin resistance disrupts: the insulin signal arrives, but the GLUT4 doors fail to open properly, so glucose stays stuck in the blood.
So already we have our first dependency: in muscle and fat, glucose entry is gated by insulin. In brain and red blood cells, it is not.
Step 2: Glycolysis — splitting glucose in the cytosol
Once inside, glucose is in the cytosol — the watery interior of the cell, outside the mitochondria. Here the first stage of breakdown happens: glycolysis (literally "sugar-splitting").
Glycolysis is a sequence of ten enzyme-driven reactions that cut the six-carbon glucose molecule into two three-carbon molecules of pyruvate. You do not need to memorise the ten steps, but a few features matter enormously:
-
It costs ATP to start. The first steps actually spend 2 ATP to prime the glucose — like priming a pump before water flows. This is why glycolysis needs a small energy investment to get going.
-
It produces a little ATP directly. Later steps generate 4 ATP, for a net gain of 2 ATP per glucose. This is a small amount — the big payoff comes much later, in the mitochondria. But it is fast, and it needs no oxygen.
-
It captures electrons onto NADH. During glycolysis, 2 molecules of NAD⁺ pick up high-energy electrons and become NADH. Hold onto this — NADH is the electron-delivery truck, and it will reappear constantly. (NAD⁺ stands for nicotinamide adenine dinucleotide; it is built from vitamin B3, niacin. When it is carrying electrons it is written NADH; when empty it is NAD⁺.)
-
The first committed step requires magnesium. The enzyme hexokinase, which traps glucose in the cell, uses ATP — and as we will see, all ATP in the body functions as a magnesium complex (Mg-ATP). No magnesium, no functional ATP.
At the end of glycolysis we have, per glucose: 2 pyruvate, a net 2 ATP, and 2 NADH. The pyruvate now faces a fork in the road.
Step 3: The pyruvate fork — with or without oxygen
Pyruvate's fate depends on whether oxygen is available and whether the mitochondria can keep up.
-
If oxygen is plentiful (the normal aerobic case), pyruvate is transported into the mitochondria for full processing. This is where the real energy is.
-
If oxygen is scarce — for example, in a muscle sprinting harder than its oxygen supply can sustain — pyruvate is instead converted to lactate (lactic acid). This is a detour, not a dead end: its purpose is to regenerate NAD⁺. Recall that glycolysis needs NAD⁺ to keep running; if all the NAD⁺ is stuck in the NADH form because the mitochondria are backed up, glycolysis would grind to a halt. Converting pyruvate to lactate hands the electrons off and frees up NAD⁺, letting glycolysis (and its quick 2-ATP-per-glucose) continue without oxygen. The lactate is later shipped to the liver and rebuilt into glucose. This is why intense exercise produces lactate — not because the muscle is "failing," but because it is running the fast anaerobic shortcut.
For the rest of Part 1 we follow the aerobic path, because that is where the overwhelming majority of energy comes from.
Step 4: Into the mitochondria — pyruvate becomes acetyl-CoA
Pyruvate crosses into the inner compartment of the mitochondrion (the matrix — the innermost space, enclosed by the folded inner membrane). There it meets one of the most important and most frequently bottlenecked enzymes in the whole body: the pyruvate dehydrogenase complex, abbreviated PDH.
PDH performs a one-way conversion: it strips a carbon off pyruvate (released as CO₂ — this is some of the carbon dioxide you breathe out), captures more electrons onto NADH, and attaches what remains to a carrier molecule called coenzyme A (CoA), producing acetyl-CoA. Acetyl-CoA is the universal "fuel-ready" molecule — the common entry token for the Krebs cycle, and as we will see, the point where fat metabolism merges with sugar metabolism.
Here is the part that matters for biohacking. PDH cannot function without a cofactor called thiamine pyrophosphate (TPP), which the body builds from vitamin B1 (thiamine). TPP is not optional — it is physically required for PDH to grip and process pyruvate. When B1 is deficient, PDH stalls. Pyruvate piles up, gets diverted into lactate, and the cell is left unable to feed the Krebs cycle efficiently from glucose. The enzyme protein is still there and intact; it is simply missing the tool it needs to do the job. This is precisely why thiamine supplementation can have such an outsized effect on energy in people who are deficient: you are not adding fuel, you are unblocking the gate that lets fuel through. (See the Thiamine deep dive for the full story, including why the brain is especially vulnerable.)
PDH is also a major regulatory checkpoint. It can be switched off by a "stop" signal (phosphorylation by an enzyme called PDH kinase) when the cell is already flush with energy or is burning fat instead — we return to this in the Randle cycle section. For now, the key idea: PDH is the gateway from sugar into the mitochondrial furnace, and it depends absolutely on vitamin B1.
Step 5: The Krebs cycle — stripping electrons off the fuel
Acetyl-CoA now enters the Krebs cycle, also called the citric acid cycle or TCA cycle (tricarboxylic acid cycle — three different names for the same loop; do not let the synonyms confuse you). It takes place in the mitochondrial matrix.
The single most important thing to understand about the Krebs cycle is what it is for. It is not primarily an ATP-making machine. Its real job is to act as a dismantling line: it takes the two carbons of acetyl-CoA and, over eight enzymatic steps arranged in a loop, completely takes them apart, releasing the carbons as CO₂ and — crucially — harvesting the high-energy electrons and loading them onto carrier molecules.
Per turn of the cycle (i.e. per acetyl-CoA), it produces:
- 3 NADH — the same electron-carrier we met in glycolysis (built from B3).
- 1 FADH₂ — a second type of electron carrier, built from vitamin B2 (riboflavin). (FADH₂ stands for flavin adenine dinucleotide, reduced form.)
- 1 GTP (which is easily converted to 1 ATP) — a small direct payoff, the cycle's only direct energy molecule.
- 2 CO₂ — waste carbon, breathed out.
Because each glucose produced two acetyl-CoA, the cycle turns twice per glucose, doubling those numbers.
Notice the pattern: the actual ATP from the Krebs cycle is trivial. What the cycle is really doing is charging up NADH and FADH₂ — loading the electron-delivery trucks. Those trucks now drive to the electron transport chain, where the real energy extraction happens.
The Krebs cycle also has its own cofactor dependencies, which is why several B vitamins and minerals show up as "energy" supplements:
- B1 (as TPP) is required again here, by an enzyme inside the cycle called α-ketoglutarate dehydrogenase (α-KGDH). This enzyme is even more sensitive to B1 deficiency than PDH, and in the brain it is often the first thing to fail.
- B2 (riboflavin) builds FAD/FADH₂.
- B3 (niacin) builds NAD⁺/NADH.
- Magnesium is required by several of the enzymes and by every ATP-using step.
- Lipoic acid is a cofactor for both PDH and α-KGDH.
A deficiency in any of these does not break one isolated reaction — it throttles the entire energy supply at once, which is why broad fatigue is such a common and non-specific symptom.
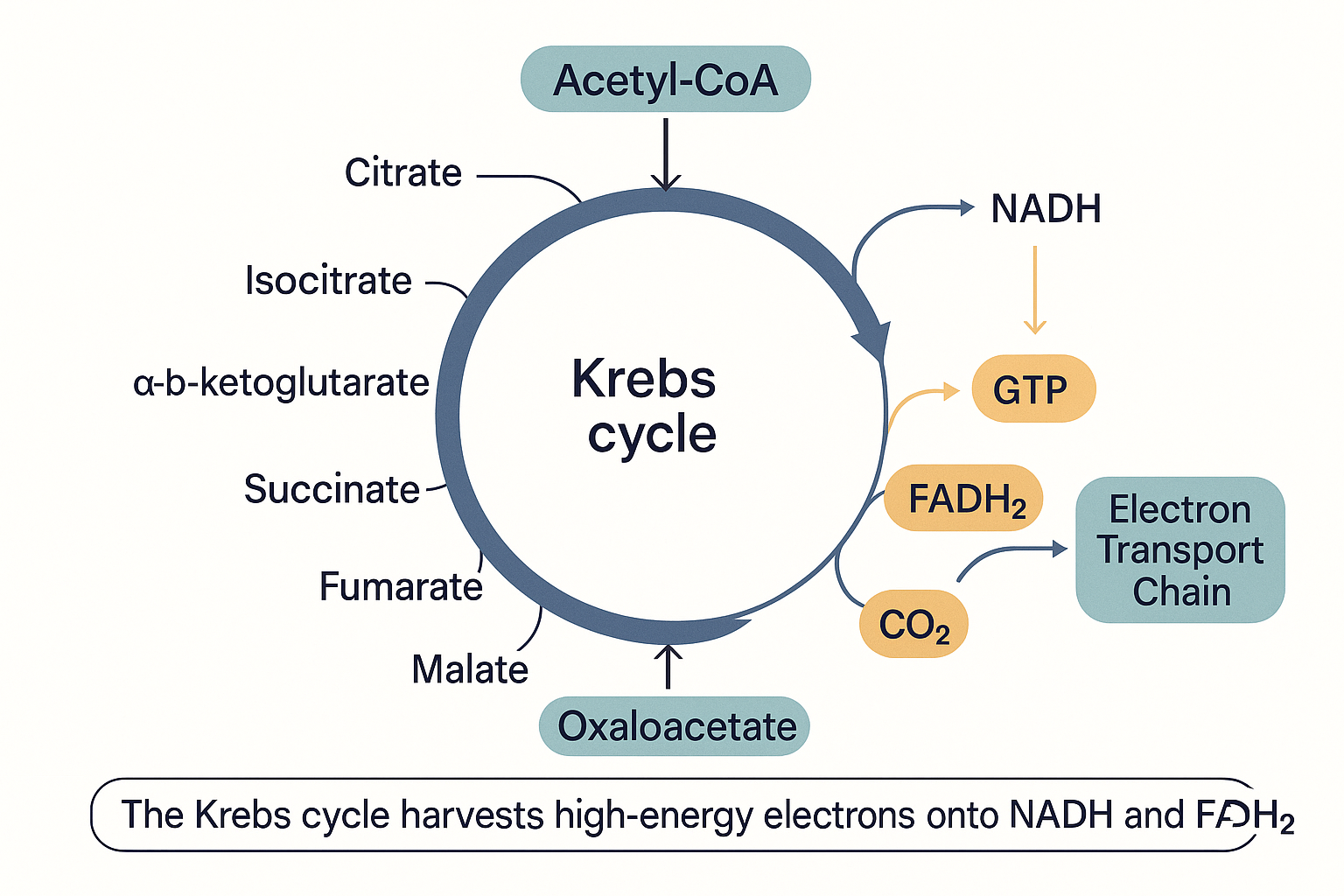 The Krebs cycle's real product is not ATP but the charged electron-carriers NADH and FADH₂, which carry the energy onward to the electron transport chain.
The Krebs cycle's real product is not ATP but the charged electron-carriers NADH and FADH₂, which carry the energy onward to the electron transport chain.
Step 6: The electron transport chain — where the energy actually comes out
We have arrived at the heart of the system. So far the cell has gained only a trickle of ATP (2 from glycolysis, 2 from the two Krebs turns). The vast majority — roughly 90% of all the ATP from a glucose molecule — is produced here, in the electron transport chain (ETC), also called the respiratory chain.
The ETC is a series of large protein structures embedded in the inner mitochondrial membrane (the heavily folded inner wall of the mitochondrion; the folds, called cristae, exist to pack in as much of this machinery as possible). There are four main complexes, numbered I to IV, plus the ATP synthase turbine (sometimes called Complex V). Here is what happens:
-
NADH delivers its electrons to Complex I. The electron-carrier trucks we charged up earlier now dock and unload. NADH drops its high-energy electrons into Complex I and reverts to empty NAD⁺ (which goes back to collect more).
-
FADH₂ delivers its electrons to Complex II. FADH₂ enters one step later, at Complex II, which is why it ultimately yields slightly less ATP than NADH — it skips the first pumping station.
-
Electrons flow "downhill" through the complexes. From Complex I or II, the electrons are passed along a chain: to a small mobile carrier called coenzyme Q10 (CoQ10, also called ubiquinone), then to Complex III, then to another mobile carrier called cytochrome c, then to Complex IV. At each major complex (I, III, IV), the energy released as electrons move to a lower energy level is used to do mechanical work: pumping protons (H⁺ ions) out of the matrix, across the inner membrane, into the space between the two mitochondrial membranes.
Note CoQ10's role here: it is the shuttle that ferries electrons between the complexes. This is exactly why CoQ10 (and its more absorbable form, ubiquinol) is taken as an energy supplement — and why the cholesterol drugs called statins, which inadvertently lower the body's CoQ10 production, can cause fatigue and muscle aches. (See CoQ10 and ubiquinol.)
- Oxygen collects the spent electrons at the end. After passing through Complex IV, the now low-energy electrons need somewhere to go, or the whole chain backs up. Their final destination is oxygen. At Complex IV, oxygen accepts the electrons and combines with protons to form water (H₂O). This is the reason you breathe. Every breath of oxygen exists to sit at the end of this chain and accept spent electrons. Without oxygen, the electrons have nowhere to go, the chain jams, NADH cannot unload, the Krebs cycle backs up behind it, and the cell is forced back onto the feeble anaerobic lactate shortcut. This is why oxygen deprivation kills energy-hungry tissue (like the brain) within minutes.
By the end of this step, the cell has pumped a large number of protons out of the matrix, creating a steep imbalance: many protons on one side of the inner membrane, few on the other. That imbalance is stored energy — and the next step cashes it in.
flowchart LR
NADH -->|electrons| CI[Complex I]
FADH2 -->|electrons| CII[Complex II]
CI --> Q[CoQ10]
CII --> Q
Q --> CIII[Complex III]
CIII --> CYC[Cytochrome c]
CYC --> CIV[Complex IV]
CIV -->|spent electrons| O2["Oxygen → water"]
CI -. pumps H+ .-> IMS[(H+ build-up<br/>outside membrane)]
CIII -. pumps H+ .-> IMS
CIV -. pumps H+ .-> IMS
IMS ==>|H+ rush back through| ATPS["ATP synthase<br/>→ ATP"]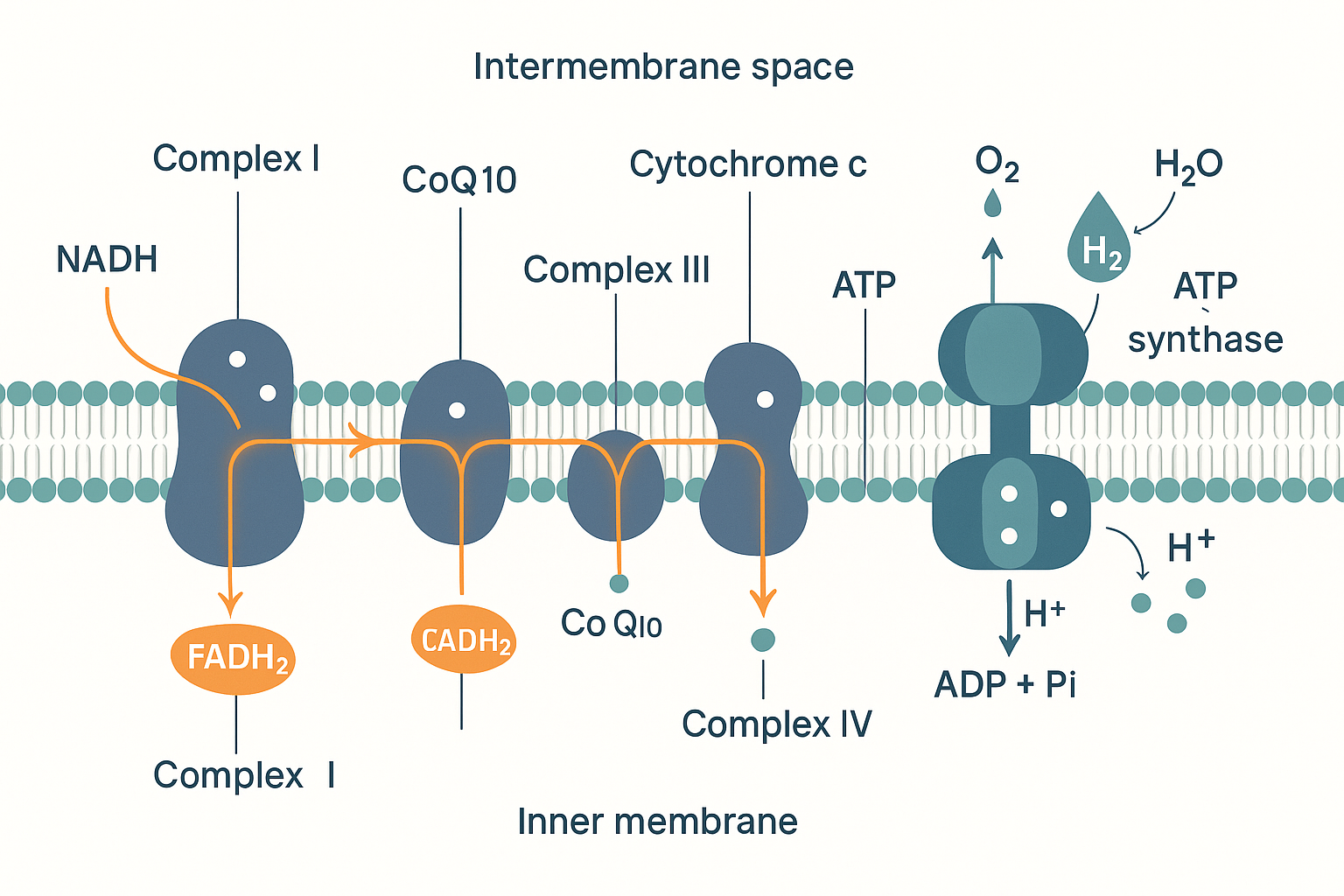 Electrons flow down the complexes; the energy released pumps protons across the membrane; those protons flow back through ATP synthase to make ATP; oxygen collects the spent electrons as water.
Electrons flow down the complexes; the energy released pumps protons across the membrane; those protons flow back through ATP synthase to make ATP; oxygen collects the spent electrons as water.
Step 7: ATP synthase — the turbine that makes the ATP
The protons pumped out in Step 6 are now crowded together on the outer side of the inner membrane. Like water held behind a dam, or charge stored in a battery, this represents potential energy. Scientists call it the proton-motive force — a combination of a concentration difference (lots of protons on one side) and an electrical charge difference across the membrane.
The protons "want" to flow back into the matrix to equalise, but the membrane blocks them — except at one specific doorway: an enzyme called ATP synthase. ATP synthase is, quite literally, a molecular turbine. As protons rush back through it, they cause part of the enzyme to physically rotate, like water spinning a hydroelectric generator. That rotation mechanically forces ADP and a phosphate group together, snapping them into ATP.
This is the elegant core of the whole system: the energy from food is not used to make ATP directly. It is first converted into a proton gradient (a pressure), and that pressure spins a turbine that makes ATP. The technical name for this mechanism is chemiosmosis (energy stored as a chemical gradient across a membrane, used to do osmotic-like work).
And here, again, magnesium: ATP synthase produces ATP, but ATP is only biologically functional as Mg-ATP — the magnesium ion stabilises the phosphate groups so that enzymes can recognise and use the molecule. This is why magnesium status quietly underlies the entire energy system: it is required at the input (glycolysis), throughout the Krebs cycle, and at the final output. (See the Magnesium deep dive.)
Step 8: The tally — how much ATP per glucose?
Adding it all up, a single glucose molecule fully processed with oxygen yields, in practice, roughly 30–32 ATP:
| Stage | Direct ATP | Electron carriers produced |
|---|---|---|
| Glycolysis (cytosol) | 2 (net) | 2 NADH |
| Pyruvate → acetyl-CoA (×2) | 0 | 2 NADH |
| Krebs cycle (×2 turns) | 2 (as GTP) | 6 NADH, 2 FADH₂ |
| Electron transport chain | ~26–28 | (consumes all the above carriers) |
| Total | ~30–32 ATP |
You sometimes see the older textbook figure of 36–38; the lower number (~30–32) reflects the real-world cost of shuttling things across membranes. The exact figure matters less than the proportion: glycolysis alone gives you 2; the mitochondria give you the other ~30. This is why mitochondrial health dominates the energy conversation. A cell that can only run glycolysis (because its mitochondria are damaged, or oxygen-starved, or missing cofactors) is operating at roughly one-fifteenth of its potential energy output.
Step 9: What is ATP actually for, and where does it go?
We have built a lot of ATP. What happens to it?
First, ATP does not travel far. It is not a hormone; it does not circulate in the blood to be delivered around the body. It is made and used locally, often within milliseconds, right where it is needed. Most ATP is produced in the mitochondrial matrix, and it gets exported into the surrounding cytosol by a dedicated swap-door protein called the adenine nucleotide translocase (ANT) — which trades a fresh ATP out of the mitochondrion for a spent ADP coming back in to be recharged. So ATP leaves the mitochondrion (into the cell body) but does not leave the cell.
What does it power? Essentially every active process in the cell:
- Muscle contraction — the proteins that pull muscle fibres together are driven by ATP. (This is also why rigor mortis sets in after death: no ATP to release the muscle proteins.)
- Pumping ions across membranes — for example the sodium-potassium pump (Na⁺/K⁺-ATPase), which every nerve and cell uses to maintain its electrical charge. This single pump consumes a startling share of the body's energy — on the order of 20–40% of resting metabolism — because it runs constantly in every cell. (This pump is also a key target of thyroid hormone — see below.)
- Building molecules — synthesising proteins, DNA, hormones, and neurotransmitters all costs ATP.
- Transport and signalling — moving cargo around the cell, firing nerve signals, secreting substances.
There is also a fast local buffer worth knowing about: the phosphocreatine system. Cells (especially muscle and brain) keep a reserve of creatine loaded with a phosphate group. When ATP is suddenly demanded faster than the mitochondria can supply, phosphocreatine instantly donates its phosphate to rebuild ATP from ADP — a rechargeable quick-release battery sitting on top of the slower main supply. This is the mechanistic basis for creatine supplementation: more phosphocreatine reserve means a larger instant-energy buffer, useful for both muscle power and, increasingly recognised, cognitive demand.
Step 10: Uncoupling — when the cell makes heat instead of ATP
Here is a beautiful twist. Remember that the proton gradient (Step 6) is what drives ATP synthase to make ATP. But what if the cell deliberately lets those protons leak back across the membrane without passing through ATP synthase?
Then no ATP is made — and the energy of the gradient is released as heat instead. This is called uncoupling: the electron transport chain keeps running and pumping protons, but it is "uncoupled" from ATP production.
The body does this on purpose. Specialised brown fat tissue contains a protein called UCP1 (uncoupling protein 1), a controlled proton leak channel. When you are cold, the body activates UCP1, the proton gradient is short-circuited, and the mitochondria pour out heat to keep you warm. This is thermogenesis — making heat from fuel. Infants have a lot of brown fat for exactly this reason; adults retain some, and cold exposure can recruit more.
This concept also explains a class of compounds. A mitochondrial uncoupler is a substance that makes the inner membrane leaky to protons, forcing the cell to burn fuel rapidly as heat rather than storing it. BAM-15 is a research compound that does exactly this in a controlled way, which is why it is investigated for fat loss. The infamous (and dangerous) drug DNP works the same way — and its lethality, from uncontrolled overheating, illustrates exactly how much energy the proton gradient holds.
Uncoupling is not just a curiosity: a baseline level of natural proton leak accounts for a meaningful fraction of your resting metabolic rate. Part of what "having a fast metabolism" means is having more uncoupling and more mitochondrial activity producing background heat.
Step 11: What controls the speed of the whole chain?
The system is demand-driven — it speeds up when energy is needed and idles when it is not. The main control signals:
-
The ADP/ATP ratio. This is the master throttle. When the cell uses ATP, it accumulates ADP. High ADP is the signal "we need more energy" — it directly accelerates the electron transport chain and ATP synthase. When ATP is abundant and ADP is low, the chain slows. The system literally runs as fast as ATP is being spent, no faster. (A related signal, AMP, builds up when energy is really low and activates a master energy sensor called AMPK, which switches the cell into fuel-burning, energy-conserving mode — the same sensor activated by exercise and by the drug metformin.)
-
The NAD⁺/NADH ratio. If NADH piles up and NAD⁺ runs short (because the electron transport chain cannot offload electrons fast enough — e.g. low oxygen), the upstream pathways (glycolysis, Krebs) stall for lack of empty NAD⁺ carriers. Keeping NAD⁺ available is so central that NAD⁺ precursors (NMN, NR, niacin) are a whole supplement category aimed at supporting it, especially as NAD⁺ levels decline with age.
-
Oxygen availability. No oxygen at the end of the chain → the whole chain backs up → fall back to anaerobic glycolysis. Oxygen is the silent dependency behind everything.
-
Substrate availability. No fuel (glucose or fat) entering, or a missing cofactor (B1, B2, B3, magnesium, CoQ10), throttles the relevant step.
-
Hormonal signals. Two are dominant. Insulin opens the glucose doors (GLUT4) and promotes fuel storage and use. Thyroid hormone (T3) sets the overall tempo of the entire system — it increases the number of mitochondria, increases the amount of electron transport chain machinery, and increases expression of the energy-consuming sodium-potassium pump and of uncoupling proteins. This is the molecular meaning of "thyroid controls your metabolic rate": it is dialling up the quantity and activity of the very machinery described on this page. (See T3 and T4; the full thyroid system gets its own foundations page.)
This is the engineer's summary: the chain runs as fast as ATP is spent (ADP), as fast as oxygen and cofactors allow, and at an overall tempo set by thyroid hormone.
Part 2 — Fat as fuel
Everything so far has followed glucose. But fat is, gram for gram, the body's largest and most energy-dense fuel store. Now we trace how fat enters the same system — and answer a question that confuses many people: can the mitochondria burn fat directly, or is it only glucose?
The answer is that mitochondria burn fat directly and extensively — in fact, at rest and during low-intensity activity, fat is the dominant fuel for most of your body. The misconception that "cells only use glucose for energy" comes from one genuine exception (the brain, discussed below), wrongly generalised to the whole body.
Step 1: Getting fat out of storage and into the blood
Body fat is stored in fat cells (adipocytes) as triglycerides — a glycerol backbone with three fatty acid chains attached. When the body needs energy and insulin is low (fasting, between meals, during exercise), hormones signal the fat cells to break triglycerides apart and release the free fatty acids into the blood. This release step is itself a major control point: insulin strongly suppresses it. This is one reason chronically high insulin (from constant eating or insulin resistance) makes it hard to access stored fat — the release valve is held shut.
Once in the blood, fatty acids do not dissolve well in water, so they travel bound to a carrier protein, albumin, which ferries them to tissues that need fuel.
Step 2: Getting fatty acids into the cell
At the target cell, the fatty acid is released from albumin and crosses the cell membrane. Because fatty acids are fat-soluble, they pass through the fatty membrane far more readily than glucose did — but the process is still assisted by transport proteins, the most important being CD36 and the FATP family (fatty acid transport proteins). The amount of CD36 a cell displays largely determines how much fat it takes up; trained muscle has more, which is part of what it means to be "fat-adapted."
Contrast this with glucose: glucose needed insulin to open GLUT4 doors. Fatty acid uptake is much less insulin-dependent and tracks mainly with supply and demand — when fatty acids are abundant in the blood and the cell needs fuel, it takes them up.
Step 3: The carnitine shuttle — the real gateway
Now the critical step, and the answer to "does fat go straight into the mitochondria?"
A fatty acid that is inside the cell is still in the cytosol — it is not yet in the mitochondrial matrix where the burning happens. And here is the catch: long-chain fatty acids cannot cross the inner mitochondrial membrane on their own. They need a dedicated ferry system, the carnitine shuttle.
It works like this:
- The fatty acid is first tagged with coenzyme A (forming fatty-acyl-CoA) in the cytosol.
- An enzyme on the outer mitochondrial membrane, carnitine palmitoyltransferase 1 (CPT1), swaps the CoA tag for a molecule of carnitine, producing fatty-acyl-carnitine.
- This carnitine-tagged fatty acid is ferried across the inner membrane by a transporter.
- On the inside, a partner enzyme, CPT2, swaps the carnitine back off for CoA, regenerating fatty-acyl-CoA now safely inside the matrix. The carnitine returns to do it again.
flowchart LR
FA["Fatty-acyl-CoA<br/>(in cytosol)"] -->|"CPT1<br/>★ the gate ★"| FAC[Fatty-acyl-carnitine]
FAC -->|crosses inner membrane| FAC2["Fatty-acyl-carnitine<br/>(in matrix)"]
FAC2 -->|CPT2 swaps carnitine back| FA2["Fatty-acyl-CoA<br/>(in matrix)"]
FA2 --> BOX[Beta-oxidation → acetyl-CoA]
MAL[malonyl-CoA<br/>made when sugar is plentiful] -. blocks .-> FA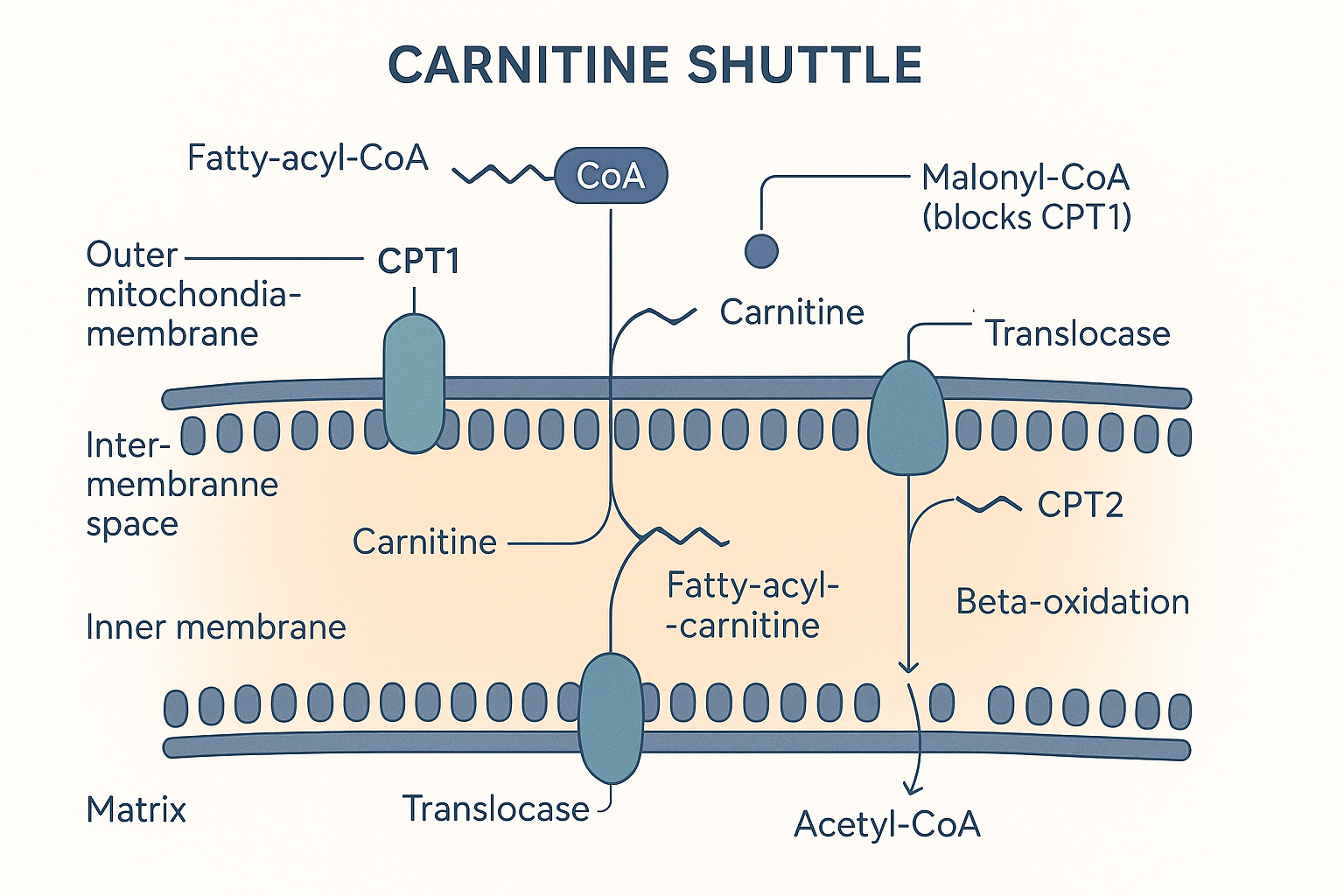 Long-chain fat cannot cross the inner membrane alone. CPT1 is the gated entry point — and malonyl-CoA, made when sugar is plentiful, holds that gate shut.
Long-chain fat cannot cross the inner membrane alone. CPT1 is the gated entry point — and malonyl-CoA, made when sugar is plentiful, holds that gate shut.
CPT1 is the single most important control point in fat burning. It is the gate, and it is deliberately regulated: when the cell is well-fed on carbohydrate, a molecule called malonyl-CoA blocks CPT1, slamming the fat-import gate shut. This is the body's logic — don't burn fat while sugar is plentiful — and it is one half of the Randle cycle discussed below.
The carnitine dependency is also why L-carnitine is supplemented for fat metabolism and endurance: carnitine is the consumable ferry token for getting long-chain fat into the furnace. (The body makes its own carnitine, so supplementation mainly helps in deficiency or specific demand states rather than acting as a simple "fat burner.")
One important exception: medium-chain fatty acids. Shorter fatty acids — medium-chain triglycerides (MCTs), found notably in coconut oil — do not need the carnitine shuttle. They are small enough to diffuse into the mitochondria directly and be burned almost immediately. This is exactly why MCT/coconut oil is prized as a "fast" fat fuel and is readily converted by the liver into ketones: it bypasses the rate-limiting CPT1 gate entirely.
Step 4: Beta-oxidation — chopping fat into acetyl-CoA
Once inside the matrix, the fatty acid undergoes beta-oxidation (β-oxidation). The name describes the mechanism: the enzyme machinery repeatedly chops two carbons off the end of the fatty acid chain, cycle after cycle, like slicing a sausage into two-carbon pieces.
Each two-carbon piece comes off as — acetyl-CoA. The exact same acetyl-CoA that glucose produced via PDH. This is the convergence point: sugar and fat, processed by completely different front-ends, both arrive at acetyl-CoA and feed the identical Krebs cycle and electron transport chain from there.
Each chop of beta-oxidation also directly produces electron carriers — 1 FADH₂ and 1 NADH per cycle — which go straight to the electron transport chain, on top of the acetyl-CoA that goes to the Krebs cycle.
This is why fat yields so much more energy than glucose per gram. A typical fatty acid (say, palmitate, 16 carbons) gets chopped into 8 acetyl-CoA, while throwing off 7 rounds' worth of FADH₂ and NADH along the way, and each of those 8 acetyl-CoA then drives a full turn of the Krebs cycle. The total comes to roughly 106 ATP from one palmitate molecule, versus ~30–32 from glucose. Fat is the high-density fuel; glucose is the fast-access fuel.
Step 5: Why the brain mostly can't use fat — and the ketone workaround
If fat is such a rich fuel, why is the brain so dependent on glucose?
Because long-chain fatty acids do not cross the blood-brain barrier — the tightly sealed wall protecting the brain from the bloodstream. Albumin-bound fatty acids simply cannot get in efficiently, so neurons cannot rely on fat the way muscle does. Under normal conditions the brain runs almost entirely on glucose, consuming about 20% of your total glucose despite being 2% of your body weight.
But there is a workaround, and it is one of the most elegant features of human metabolism: ketone bodies. When glucose is scarce (fasting, prolonged low-carb intake), the liver takes the flood of acetyl-CoA it is generating from heavy fat breakdown and packages it into small, water-soluble molecules called ketones (beta-hydroxybutyrate and acetoacetate). Ketones can cross the blood-brain barrier. The brain takes them up, converts them back into acetyl-CoA, and feeds them into its own Krebs cycle. In effect, ketones are the liver's way of converting fat into a brain-usable fuel — a fat-derived currency that can reach the one organ fat itself cannot.
This is the mechanistic basis of ketogenic diets and fasting: drive insulin down, release stored fat, flood the liver with acetyl-CoA, and produce ketones that fuel the brain in glucose's place. Note the dependency chain — it only works when insulin is low enough to permit large-scale fat release in the first place.
Step 6: The Randle cycle — glucose and fat compete
The final piece ties Part 1 and Part 2 together. The cell does not burn glucose and fat in fixed proportions; it switches between them, and the two fuels actively suppress each other. This reciprocal relationship is the Randle cycle (or glucose–fatty-acid cycle).
- When fat is being burned heavily, beta-oxidation floods the cell with acetyl-CoA and NADH. These build up and send "stop" signals to PDH — switching off the entry of glucose-derived pyruvate into the mitochondria. The message: we're running on fat, don't also pour in sugar.
- When glucose is being burned heavily (after a carbohydrate meal, insulin high), the cell produces malonyl-CoA, which blocks CPT1 — slamming shut the fat-import gate from Part 2 Step 3. The message: we're running on sugar, stop importing fat.
So the two pathways are wired as a see-saw, each one's activity inhibiting the other's entry point.
flowchart LR
GLU["Burning glucose<br/>(↑ malonyl-CoA)"] -. blocks CPT1 .-> NOFAT[Fat import shut off]
FATB["Burning fat<br/>(↑ acetyl-CoA + NADH)"] -. blocks PDH .-> NOGLU[Glucose entry shut off]This is why metabolism has a "fuel selection" behaviour: fed and carbohydrate-rich tilts toward glucose-burning and fat-storage; fasted and low-insulin tilts toward fat-burning and ketone production. And it is why metabolic flexibility — the ability to switch cleanly between the two — is considered a marker of metabolic health, while being "stuck" (unable to access fat because insulin is chronically high, or unable to use glucose well because of insulin resistance) is a marker of dysfunction.
Putting it all together
Step back and look at the whole engine:
- Fuel enters the cell — glucose through insulin-gated GLUT doors, fat through CD36 transporters.
- Front-end processing differs — glucose runs glycolysis in the cytosol; fat runs beta-oxidation in the matrix after the carnitine shuttle — but both converge at acetyl-CoA.
- The Krebs cycle dismantles acetyl-CoA, not to make ATP, but to charge up the electron carriers NADH and FADH₂.
- The electron transport chain discharges those carriers, using their electrons to pump protons and build a gradient, with oxygen collecting the spent electrons at the end.
- ATP synthase lets the protons flow back through it and spins out the ATP — about 90% of the total.
- ATP is spent locally — never circulating, recharged millions of times a second, buffered by phosphocreatine, powering pumps and contraction and synthesis.
- The whole system is throttled by ADP demand, NAD⁺ supply, oxygen, cofactors, and — for overall tempo — thyroid hormone, while glucose and fat trade off against each other via the Randle cycle.
Every cofactor that appears as an "energy supplement" maps onto a specific point in this chain: B1 at the PDH and α-KGDH gates, B2 building FADH₂, B3 building NADH, magnesium at every ATP step, CoQ10 ferrying electrons mid-chain, carnitine ferrying fat in, creatine buffering the output. None of them is an energy source. Every one of them is a tool that removes a bottleneck — which is exactly why they only help when that particular bottleneck is the thing holding you back.
Related Compounds & Deep Dives
These pages connect directly to the machinery described above:
Cofactors and carriers in the chain
- Thiamine (B1) deep dive — TPP is the required cofactor for PDH and α-KGDH, the two gateway enzymes feeding the Krebs cycle. The clearest example of "unblocking a gate" rather than adding fuel.
- Magnesium deep dive — every functional ATP is Mg-ATP; magnesium is required at glycolysis, throughout the Krebs cycle, and at ATP synthase.
- CoQ10 / ubiquinone and ubiquinol — the mobile electron shuttle between Complexes I/II and III in the electron transport chain.
- Niacin (B3), niacinamide, NMN, NR, NAD+ — all support the NAD⁺/NADH electron-carrier pool that runs the entire chain.
- PQQ — implicated in mitochondrial biogenesis (making more mitochondria).
- TTFD and benfotiamine — fat-soluble B1 forms; see the thiamine deep dive.
Fuel handling
- L-carnitine / carnitine — the ferry token for the CPT1 carnitine shuttle that imports long-chain fat into the mitochondria.
- Coconut oil / MCTs — medium-chain fats that bypass the carnitine shuttle and are rapidly burned or turned into ketones.
- Creatine — the phosphocreatine quick-release buffer that instantly recharges ATP from ADP.
System tempo and regulation
- T3 and T4 — thyroid hormone sets the overall metabolic tempo by increasing mitochondrial number, ETC machinery, and the energy-consuming sodium-potassium pump.
- Insulin — gates glucose entry via GLUT4 and suppresses fat release; the master switch in the Randle fuel-selection see-saw.
- BAM-15 — a mitochondrial uncoupler that dissipates the proton gradient as heat instead of ATP.