Inflammation
The most over-used word in health — "inflammation" gets blamed for everything. This page makes it precise: what inflammation actually is, the exact molecular triggers that switch it on, what it looks like in the tissue, why it makes you feel terrible, why it is also one of the things keeping you alive, and why the modern version that smoulders for years is so destructive.
Why this page exists
By this point in the Foundations series, "inflammation" has appeared on nearly every page — driving insulin resistance on the metabolism page, leaking from the gut on the microbiome page, inflaming the brain on the gut-brain page, suppressing testosterone on the testosterone deep dive. It is the connective tissue of the whole series. But we have been using the word without ever defining it properly. This page fixes that.
The problem with "inflammation" in popular health talk is that it is used as a vague villain — a word that means "the bad thing" without any mechanism. The reality is precise and, once you see it, genuinely illuminating: inflammation is a specific, evolved biological program — the immune system's coordinated response to threat and damage — and almost everything confusing about it dissolves once you understand that the same program that saves your life in an acute emergency destroys your health when it runs continuously. Acute inflammation is a hero; chronic inflammation is that same hero unable to stand down. Holding those two apart is the key to the whole topic.
We build it from the definition up.
What inflammation actually is
Inflammation is the body's response to harm. More precisely: it is the coordinated reaction of the immune system (the body's defence network of cells and signals) to anything it perceives as a threat — an infection, an injury, a toxin, or damaged tissue. Its purpose is threefold: (1) neutralise the threat (kill invaders), (2) clear the debris (remove dead cells and damage), and (3) initiate repair (rebuild the tissue).
That is the whole thing. Inflammation is not a disease — it is a defence and repair operation. The redness around a cut, the swelling of a sprained ankle, the fever of a flu, the soreness of a sore throat — these are not the infection or the injury itself. They are your body's response to it. The symptoms you feel are largely the inflammation, not the threat.
The critical distinction, which the rest of the page hangs on:
- Acute inflammation is fast (minutes to days), intense, localised, and — crucially — self-terminating. It flares up, does its job, and shuts down. This is the good kind, the kind you cannot live without.
- Chronic inflammation is slow, low-grade, often body-wide, and fails to shut down — smouldering for months or years. This is the kind implicated in nearly every modern chronic disease.
Same machinery, opposite consequences. Now let us watch the acute version happen.
What acute inflammation looks like — the classic response
Cut your finger, and within minutes a recognisable sequence unfolds. The Romans named its outward signs two thousand years ago, and they still hold — the five cardinal signs: redness, heat, swelling, pain, and loss of function. Each one is the visible result of a specific mechanism:
- Redness and heat come from vasodilation — the small blood vessels at the site widen, flooding the area with warm, oxygen- and cell-rich blood. More blood = redder and warmer.
- Swelling comes from increased vascular permeability — the vessel walls become deliberately leaky, letting fluid and immune proteins seep out of the blood into the tissue. That fluid is the swelling, and it carries the molecular tools of defence to where they're needed.
- Pain comes from inflammatory molecules (notably prostaglandins and bradykinin) sensitising the local nerve endings — which is useful: pain makes you protect and rest the injured part.
- Loss of function is the sum — a swollen, painful part is one you stop using, which enforces the rest that aids healing.
Underneath those signs, a cellular operation is running. The immune cells arrive in waves:
- Neutrophils are the first responders — they swarm the site within hours, engulfing and destroying bacteria (and dying in the process; pus is largely spent neutrophils).
- Macrophages (literally "big eaters") arrive next — they devour pathogens and debris, and they orchestrate the later phases, deciding when to escalate and when to begin repair.
- Mast cells release histamine, a major driver of the vasodilation and leakiness (this is the same histamine that antihistamines block — and why allergic reactions, which are histamine-driven, look like inflammation).
So the visible flare is the surface of a precise, coordinated defence: widen the vessels, make them leaky, pour in fluid and cells, kill the threat, clear the mess. Now — what switches all this on?
flowchart TD
DAMAGE[Injury / infection at a site] --> VASO[Vessels widen<br/>vasodilation]
DAMAGE --> PERM[Vessels turn leaky<br/>↑ permeability]
VASO --> R1["Redness + heat<br/>(more warm blood)"]
PERM --> R2["Swelling<br/>(fluid into tissue)"]
PERM --> CELLS[Immune cells pour in:<br/>neutrophils → macrophages]
CELLS --> KILL[Kill pathogens · eat debris]
KILL --> REPAIR[Macrophages switch to<br/>cleanup + repair mode]
DAMAGE --> PAIN["Pain + loss of function<br/>(prostaglandins sensitise nerves)"]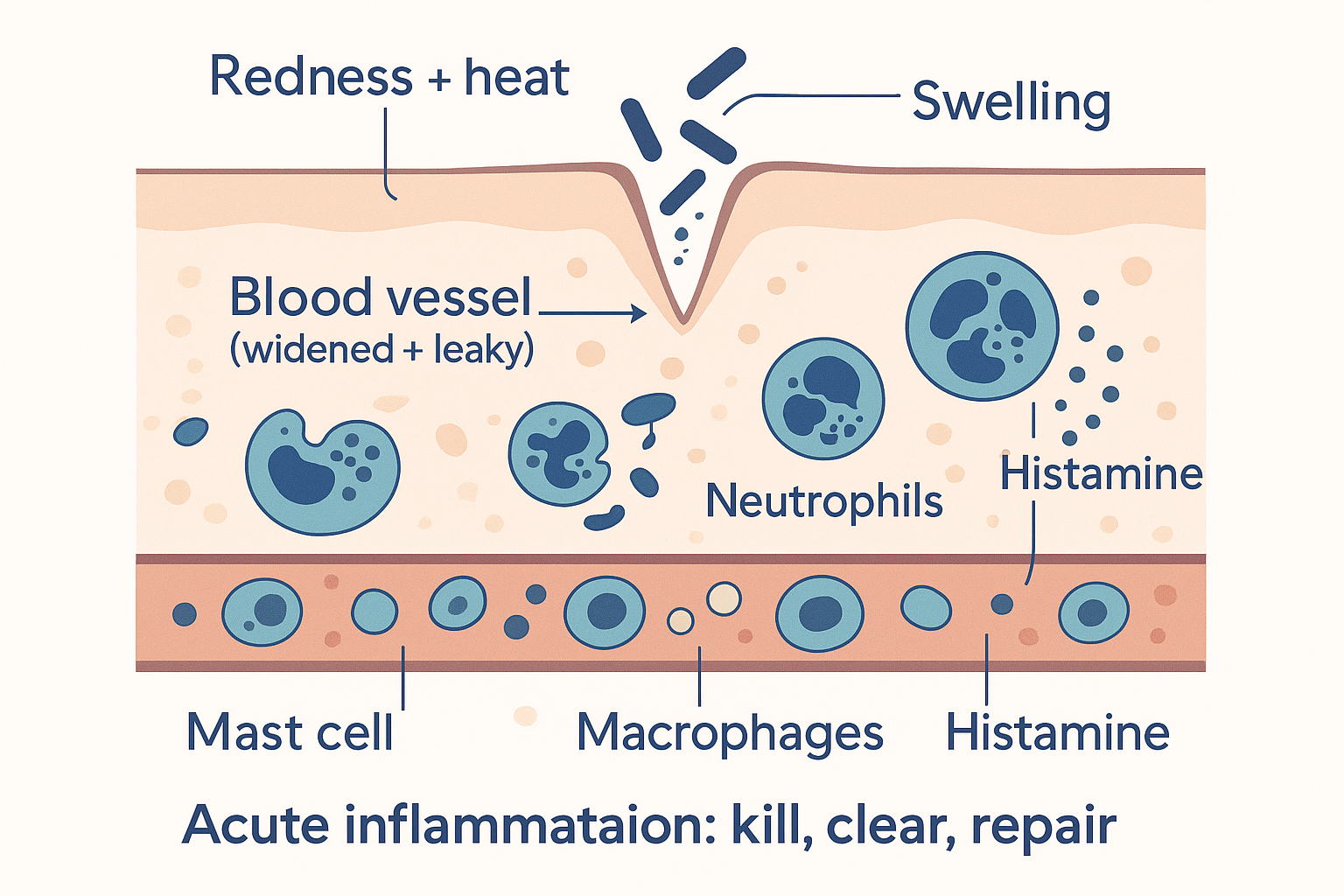 Acute inflammation in action: vessels widen and leak, immune cells pour in to kill the threat and clear debris, then switch to repair — the visible redness, heat, and swelling are the surface of that operation.
Acute inflammation in action: vessels widen and leak, immune cells pour in to kill the threat and clear debris, then switch to repair — the visible redness, heat, and swelling are the surface of that operation.
The triggers: how inflammation is switched on
This is the part that turns "inflammation" from a vague word into a mechanism. The immune system does not react randomly; it has molecular sensors that detect specific danger signals and flip a master switch. There are two broad categories of danger signal, and they are worth knowing by name because they appear across the whole Foundations series:
- PAMPs — pathogen-associated molecular patterns. These are molecules that are characteristic of invaders — bits of bacteria, viruses, fungi. The single most important example on this site is LPS (lipopolysaccharide / endotoxin), the fragment of gram-negative bacterial wall from the gut microbiome page. PAMPs scream "something foreign is here."
- DAMPs — damage-associated molecular patterns. These are molecules released by your own cells when they are injured or dying — things that should be safely inside a cell and have no business being outside it. When the immune system detects them in the open, it reads "tissue is being damaged here." This is how inflammation responds to injury and stress even without any infection.
These danger signals are detected by pattern-recognition receptors on immune cells — the most famous being the Toll-like receptors (TLRs), especially TLR4, the LPS sensor we met on the gut and testosterone pages. When a TLR catches its danger signal, it triggers an internal relay that activates the master inflammatory switch:
NF-κB (nuclear factor kappa B) — the central on-switch for inflammation. When NF-κB is activated, it enters the cell nucleus and turns on the genes for the inflammatory messenger molecules. Almost every inflammatory trigger ultimately converges on flipping NF-κB.
What NF-κB produces are the cytokines — the hormone-like signalling proteins of the immune system, the molecules that actually carry out and spread inflammation. The big three to know:
- TNF-α (tumour necrosis factor alpha) — a master inflammatory cytokine.
- IL-1β (interleukin-1 beta) — drives fever and the alarm response.
- IL-6 (interleukin-6) — spreads the signal body-wide and triggers the liver's acute-phase response.
There is also a second, parallel chemical arm worth knowing because it connects to the diet debates on other pages: the eicosanoids. These are inflammatory signalling molecules built from fatty acids in your cell membranes — specifically from arachidonic acid, a polyunsaturated fat. Two enzymes, COX (cyclooxygenase) and LOX (lipoxygenase), convert arachidonic acid into prostaglandins and leukotrienes — the molecules that cause much of the pain, fever, and vessel changes. This is mechanistically important: it is why aspirin and ibuprofen work (they block COX, cutting prostaglandin production), and it is the kernel of truth behind the PUFA debates — the polyunsaturated fats in your membranes are the literal raw material for one arm of inflammation.
flowchart TD
PAMP["PAMPs<br/>(invaders, e.g. LPS endotoxin)"] --> TLR[Pattern-recognition<br/>receptors e.g. TLR4]
DAMP["DAMPs<br/>(your own damaged cells)"] --> TLR
TLR --> NFKB["NF-κB<br/>(master ON switch)"]
NFKB --> CYTO["Cytokines<br/>TNF-α · IL-1β · IL-6"]
PUFA["Arachidonic acid<br/>(membrane PUFA)"] -->|COX / LOX| EICO["Prostaglandins<br/>+ leukotrienes"]
CYTO --> RESP["Vasodilation · leaky vessels ·<br/>immune cells recruited"]
EICO --> RESP
RESP --> ACT["Kill threat · clear debris · start repair"]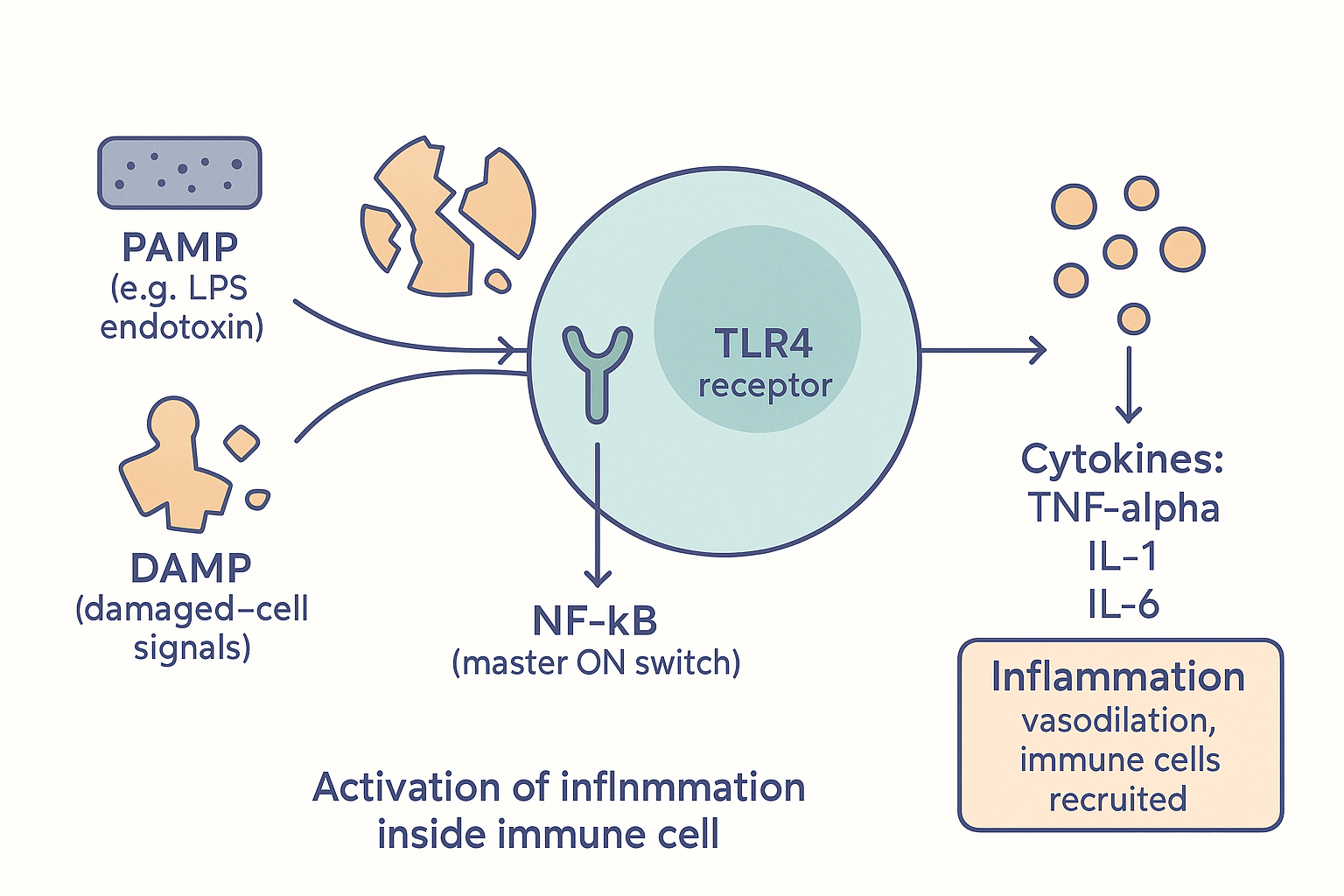 The switch: danger signals (PAMPs/DAMPs) are caught by receptors like TLR4, which flip the master switch NF-κB to produce the cytokines that carry out inflammation.
The switch: danger signals (PAMPs/DAMPs) are caught by receptors like TLR4, which flip the master switch NF-κB to produce the cytokines that carry out inflammation.
Why inflammation makes you feel terrible
Here is something most people never realise: the awful feeling of being ill — the fatigue, aches, fever, brain fog, low mood, and loss of appetite — is not caused by the infection. It is caused by your own inflammation, on purpose. This cluster has a name: sickness behaviour, and it is an evolved program, not a malfunction.
The mechanism: the cytokines (especially IL-1β, IL-6, and TNF-α) don't stay at the infection site — they circulate, and they signal the brain. The brain reads "the body is fighting something" and deliberately generates the sick feeling to change your behaviour in ways that help you win the fight:
- Fever — IL-1β acts on the hypothalamus (via prostaglandins) to raise the body's temperature set-point. A higher temperature hampers many pathogens and speeds immune cells. (This is also why aspirin/ibuprofen reduce fever — they block the prostaglandin step.)
- Fatigue and the urge to rest — fighting an infection is metabolically expensive; the brain enforces rest to redirect energy to the immune system.
- Loss of appetite — conserves energy and, by lowering available nutrients, may starve pathogens.
- Aches and pain — prostaglandins sensitise nerves; the discomfort makes you rest.
- Low mood, social withdrawal, brain fog — the cytokines act on mood and cognitive circuits (the gut-brain and cognition pages: inflammation drives the tryptophan→kynurenine diversion, lowering serotonin and raising excitotoxic metabolites). Withdrawing from others historically also limited spread of infection.
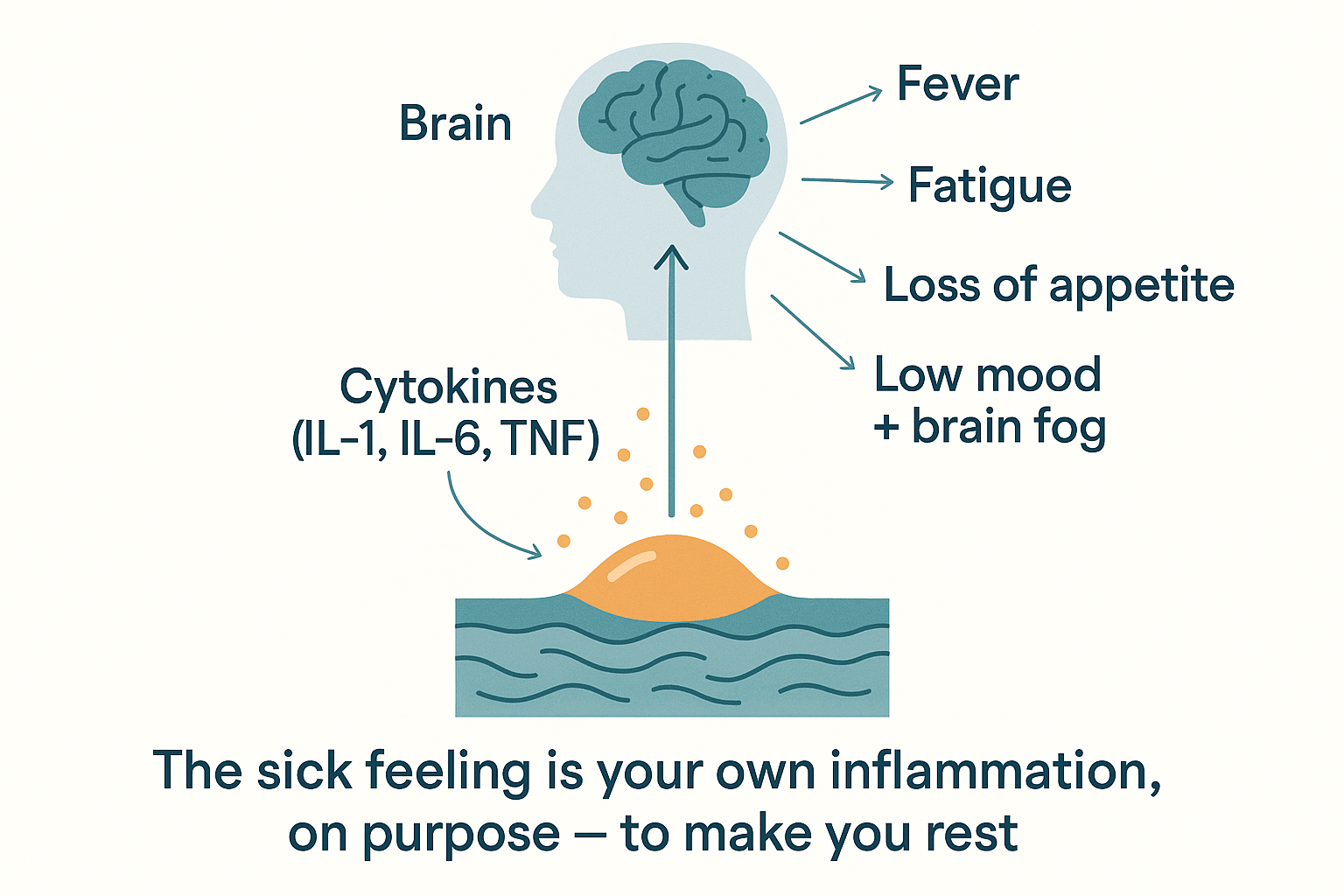 Sickness behaviour: the cytokines signal the brain, which deliberately generates the awful sick feeling to make you rest and let the immune system win.
Sickness behaviour: the cytokines signal the brain, which deliberately generates the awful sick feeling to make you rest and let the immune system win.
So feeling rotten when ill is your body deliberately making you rest and withdraw so it can win the fight. It is adaptive — for an acute infection. The trouble, as we'll see, is that when inflammation becomes chronic, this same sickness-behaviour program runs at a low level continuously — producing the persistent fatigue, brain fog, and low mood that are the everyday face of chronic inflammation. The depression-and-inflammation link is, in large part, chronic sickness behaviour.
When and why inflammation is essential
Before condemning inflammation, the other half of the truth: acute inflammation is indispensable. You would die quickly without it. Its usefulness is concrete:
- It kills pathogens. The neutrophils and macrophages that swarm a wound are the reason a minor cut doesn't become a fatal infection. People who cannot mount inflammation (e.g. certain immune deficiencies, or patients on strong immunosuppressants) are gravely vulnerable to ordinary infections.
- It clears damage. Macrophages eat dead cells and debris — the necessary cleanup before tissue can be rebuilt.
- It initiates repair. Inflammation isn't just destruction; it actively signals the start of healing. Macrophages, having cleared the site, switch phenotype and release growth and repair signals that summon the cells that rebuild tissue and blood vessels. No inflammation, no healing. A wound that cannot inflame cannot repair.
This is why "anti-inflammatory" is not a synonym for "good." Blunting all inflammation indiscriminately impairs your defence against infection and your ability to heal. The goal is never zero inflammation — it is appropriate inflammation that resolves cleanly. Which brings us to the most underrated part of the whole story.
Resolution: inflammation is supposed to end
Here is the concept that reframes everything and explains why chronic inflammation is a distinct beast. For decades, inflammation was thought to simply fade away once the trigger was gone — a passive winding-down. We now know that is wrong. Ending inflammation is an active, programmed process in its own right, called resolution.
When acute inflammation has done its job, the body actively switches it off by producing a distinct class of signalling molecules — the specialised pro-resolving mediators (SPMs): resolvins, protectins, and lipoxins. Strikingly, many of these are built from omega-3 fatty acids (and some from omega-6). They actively:
- Stop new immune cells arriving.
- Tell the neutrophils to die off and be cleared.
- Switch macrophages from "attack" mode to "cleanup and repair" mode.
- Restore the tissue to its calm baseline.
This matters enormously, because it means inflammation has a built-in off-switch that can itself fail or be under-supplied. Chronic inflammation is, in large part, not "too much triggering" but failed resolution — the inflammation never receives, or cannot execute, the signal to stand down. And one reason this off-switch may be under-resourced is a shortage of the omega-3 raw materials the resolving molecules are built from — the genuine mechanistic basis for omega-3 (fish oil) being "anti-inflammatory." It doesn't block inflammation so much as supply the materials to resolve it.
flowchart LR
TRIG[Trigger: infection / injury] --> ACUTE[Acute inflammation<br/>kill · clear · repair]
ACUTE --> RESOLVE{Resolution program<br/>SPMs: resolvins, lipoxins}
RESOLVE -->|"works → off-switch fires"| HEAL[Tissue heals,<br/>returns to baseline ✓]
RESOLVE -->|"fails / trigger persists"| CHRONIC[CHRONIC inflammation<br/>smoulders for months–years ✗]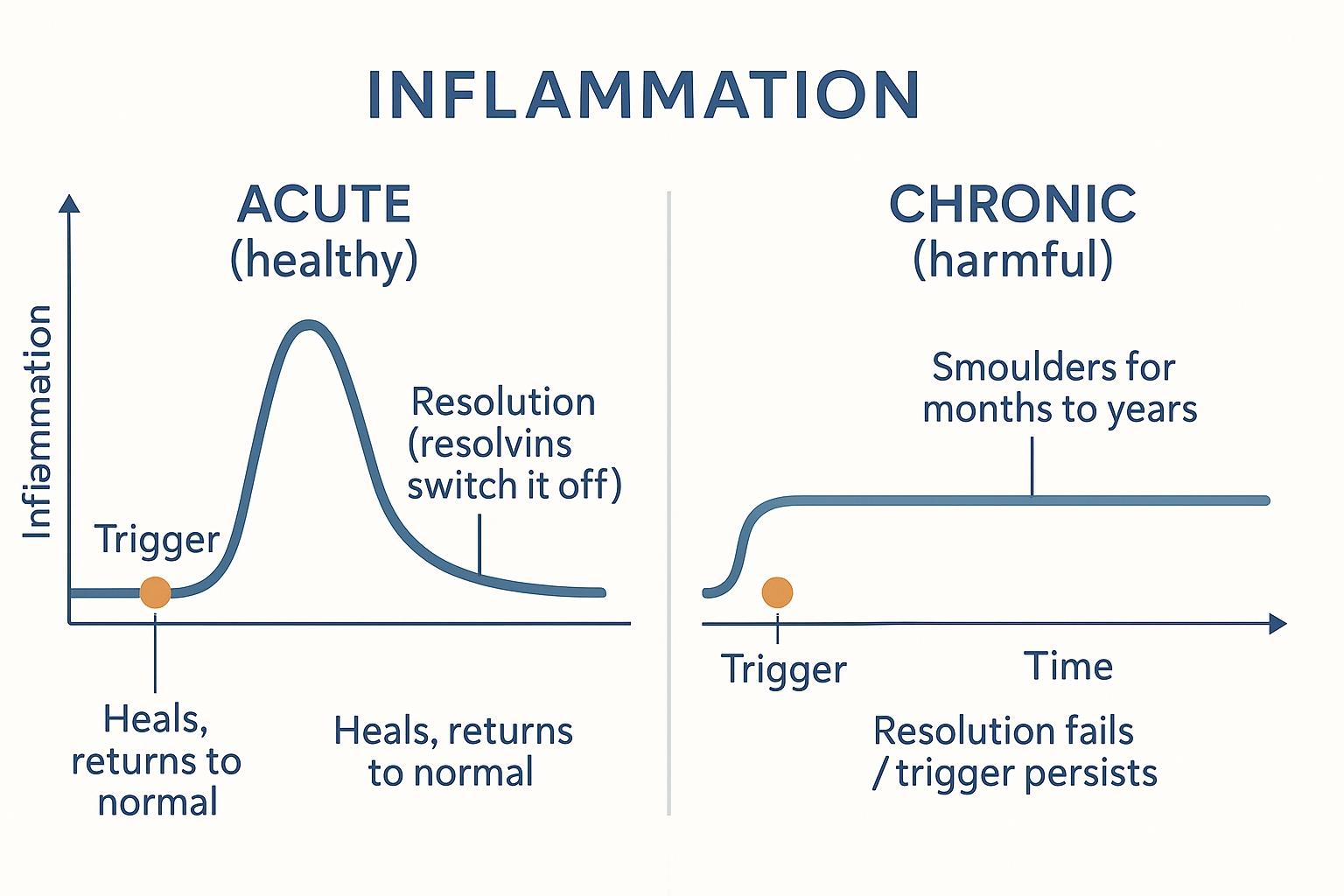 The whole story in two curves: acute inflammation spikes and is actively switched off; chronic inflammation is the same fire that never receives — or cannot execute — the signal to go out.
The whole story in two curves: acute inflammation spikes and is actively switched off; chronic inflammation is the same fire that never receives — or cannot execute — the signal to go out.
Chronic inflammation: the same fire that won't go out
Chronic inflammation is acute inflammation's machinery running at a low level, continuously, and body-wide — never fully resolving. It is sometimes called "smouldering" or "low-grade" inflammation, and a related ageing version is nicknamed "inflammaging." It usually produces no obvious redness or swelling — you can't see it — but its cytokines circulate persistently at modestly elevated levels, and over years that does enormous damage.
Why does it happen — why doesn't it resolve? Two reasons, often together:
- The trigger never goes away. Acute inflammation is built for a threat that gets dealt with and disappears. But chronic triggers persist indefinitely: a continuously leaky gut dripping LPS into the blood (microbiome page), visceral fat (which is not inert — fat cells and the macrophages within them actively secrete inflammatory cytokines, making body fat an inflammation-producing organ), a chronic low-grade infection, an autoimmune process (the immune system mistaking self for threat), ongoing oxidative stress from PUFA/poor diet, or chronic psychological stress (which, via cortisol dysregulation, fails to restrain inflammation properly). The fire keeps being re-lit.
- Resolution fails. Even without a persistent trigger, if the pro-resolving program is under-supplied (e.g. low omega-3, poor metabolic health), inflammation doesn't get switched off cleanly.
Why is it bad — why does it cause so much disease? Because the inflammatory program is destructive by design (it's meant to kill and break down), and running it continuously turns those tools against your own tissues:
- It damages tissues directly — chronic low-grade attack on blood vessel walls is central to atherosclerosis (heart disease); on joints, to arthritis; on the brain, to neurodegeneration.
- It causes insulin resistance — TNF-α and IL-6 jam the insulin signal (the exact IRS-1 mechanism from the metabolism page), driving type 2 diabetes and metabolic syndrome.
- It suppresses other systems — chronic inflammation blunts thyroid conversion, suppresses testosterone (the testosterone deep dive), and disrupts mood and cognition (the chronic sickness behaviour above — persistent fatigue, brain fog, low mood).
- It promotes cancer — a chronically inflamed environment supplies growth signals and DNA-damaging oxidants that favour tumour development.
flowchart LR
subgraph DRIVERS["Persistent triggers (the fire keeps re-lighting)"]
D1[Leaky gut → LPS endotoxin]
D2[Visceral fat<br/>secretes cytokines]
D3[Chronic stress / poor sleep]
D4[PUFA / oxidative stress]
D5[Low omega-3 → failed resolution]
end
DRIVERS --> CHRONIC[Chronic low-grade<br/>inflammation]
CHRONIC --> C1[Atherosclerosis]
CHRONIC --> C2[Insulin resistance]
CHRONIC --> C3[Neurodegeneration,<br/>brain fog, low mood]
CHRONIC --> C4[Suppressed thyroid<br/>+ testosterone]
CHRONIC --> C5[Cancer promotion]This is why chronic inflammation shows up as the common thread behind heart disease, diabetes, depression, autoimmune conditions, neurodegeneration, and ageing itself. It is not that inflammation "causes everything" in a hand-wavy way — it is that the specific destructive molecules of an unresolved immune response, applied continuously to your own tissues, degrade many systems at once.
Why does chronic inflammation even exist — why would evolution permit it? Because the acute program was never designed to run continuously — it is a mismatch. The system evolved for a world of acute threats: get injured or infected, mount a fierce local response, win, resolve, return to baseline. It is superb at that. What it was never built for is the modern environment of persistent low-grade triggers — a permanently leaky gut from processed food, chronically elevated visceral fat, relentless psychological stress, sleep deprivation, and a diet that under-supplies the omega-3s needed for resolution. Evolution optimised the acute response and assumed triggers would be temporary. Modern life supplies triggers that never stop, so the off-switch never fires. Chronic inflammation is largely an evolutionary mismatch: a brilliant emergency system trapped in a permanent emergency that isn't really one.
How chronic inflammation is lowered (in principle)
Because the cause is persistent triggers + failed resolution, the levers follow directly — and they're the same ones the rest of the Foundations series keeps arriving at:
- Remove the persistent triggers: heal the gut barrier to cut endotoxin (microbiome page), lose visceral fat (less inflammatory adipose tissue), treat chronic infections, manage stress and sleep (ANS page — vagal tone is directly anti-inflammatory).
- Support resolution: supply omega-3s (fish oil) as raw material for resolvins; the vagus nerve's cholinergic anti-inflammatory reflex is a literal neural off-switch you can strengthen with breathwork and meditation.
- Targeted anti-inflammatory tools — from aspirin (COX inhibition) to curcumin and others — but with the caveat from the "resolution" section: the aim is to support appropriate resolution, not to abolish all inflammation, which would impair defence and healing.
The single most important reframe: the goal is not to eliminate inflammation but to keep it acute, appropriate, and self-resolving — a fire you can light when needed and that reliably goes out, rather than a pilot light that never switches off.
Putting it all together
- Inflammation is the immune system's coordinated response to threat or damage — neutralise, clear, repair. The symptoms you feel are the response, not the threat.
- Acute inflammation is fast, local, and self-terminating — the classic redness/heat/swelling/pain from vasodilation, leaky vessels, and waves of immune cells (neutrophils, macrophages, mast-cell histamine). It is essential — it kills pathogens, clears debris, and initiates healing; you cannot live or heal without it.
- It is switched on when danger signals — PAMPs (invaders, e.g. LPS) and DAMPs (your own damaged cells) — are caught by pattern-recognition receptors (TLR4), flipping the master switch NF-κB, which produces the cytokines (TNF-α, IL-1β, IL-6); a parallel arm builds prostaglandins/leukotrienes from membrane PUFA (the target of aspirin/ibuprofen).
- It makes you feel terrible on purpose — circulating cytokines trigger sickness behaviour (fever, fatigue, aches, appetite loss, low mood) to make you rest and let the immune system win. Chronic low-grade inflammation runs this program continuously — the basis of inflammation-linked fatigue and depression.
- Resolution is an active program (resolvins, lipoxins, largely from omega-3) that switches inflammation off. Chronic inflammation is largely failed resolution plus persistent triggers.
- Chronic inflammation is the same machinery smouldering body-wide for years — driven by leaky-gut endotoxin, visceral fat, chronic stress, poor sleep, and under-supplied resolution. It is destructive because the inflammatory tools, applied continuously to your own tissues, cause atherosclerosis, insulin resistance, neurodegeneration, hormonal suppression, and cancer promotion.
- It exists because of evolutionary mismatch — a system built for temporary acute threats, trapped by modern persistent low-grade triggers so the off-switch never fires.
The unifying idea: inflammation is a tool, not a villain — the question is always acute-and-resolving versus chronic-and-smouldering. Nearly every "anti-inflammatory" intervention worth doing works by either removing a persistent trigger or supporting the resolution program — not by abolishing the fire, but by making sure it goes out.
Related Compounds & Deep Dives
Resolution & omega-3
- Fish oil — supplies the omega-3 raw material for resolvins and protectins, the pro-resolving mediators that switch inflammation off.
COX / eicosanoid inhibitors
- Aspirin — blocks COX, cutting prostaglandin production (and, intriguingly, triggers some pro-resolving mediators).
- Boswellia — inhibits the LOX/leukotriene arm.
- Black seed oil — broad anti-inflammatory and immune-modulating effects.
NF-κB / cytokine modulators
- Curcumin — inhibits NF-κB, the master inflammatory switch.
- Quercetin — mast-cell stabiliser (lowers histamine) and NF-κB modulator.
- EGCG — green-tea polyphenol with NF-κB-modulating activity.
- PEA (palmitoylethanolamide) — modulates mast-cell-driven inflammation.
Antioxidant / oxidative-stress support
- NAC & glutathione — counter the oxidative stress that drives and sustains inflammation.
- Vitamin E — protects membrane fats from peroxidation.
- Vitamin D — broadly immunomodulatory; low levels track higher inflammation.
Barrier & trigger removal
- Butyrate, lactoferrin — seal the gut barrier, cutting the endotoxin that is a leading chronic trigger.
- Glycine, taurine — anti-inflammatory amino acids.
Histamine / mast cell
- Histamine — the mast-cell mediator behind much acute vasodilation and the allergic overlap with inflammation.
Related foundations
- Gut Microbiome — LPS endotoxin, the archetypal chronic inflammatory trigger.
- Systemic Metabolism — how inflammation causes insulin resistance.
- Autonomic Nervous System — the vagal cholinergic anti-inflammatory reflex, a neural off-switch.
- Gut-Brain Axis — how inflammation reaches the brain to cause sickness behaviour and low mood.
- The Testosterone Kabbalah — inflammation as a suppressor of testosterone.