Systemic Metabolism: Thyroid, Hormones, and Regulatory Loops
Zoom out from the single cell to the whole body. How the thyroid sets your metabolic tempo, how insulin and cortisol partition your fuel, how hunger is regulated — and how all of these wire together into one self-regulating (and sometimes self-sabotaging) system.
Why this page exists
The cellular energy page showed how one cell turns fuel into ATP. The liver page showed how the body's central depot stores, releases, and transforms that fuel. But neither answered the body-wide question: what decides whether the whole system runs hot or cold, stores or burns, builds or breaks down? That is the job of the endocrine (hormone) system, and this page is the wiring diagram.
This is the page the user asked for explicitly: to visualise the system like an engineer — the dependencies, the feedback loops, how each part regulates the others. So we will build it that way. We introduce each major hormone as a control signal, explain what it senses and what it does, and then — the important part — show how they are wired together. Because the single most important truth about metabolism is that no hormone acts alone. Chronic stress does not just raise cortisol; it suppresses thyroid, drives insulin resistance, and disrupts hunger signalling, all at once. Understanding the loops is the whole point.
We start by clearing up what "metabolism" even means.
What "metabolism" actually means
In everyday speech "metabolism" means "how many calories you burn," and "fast metabolism" means "can eat without gaining weight." That is a shadow of the real concept. Metabolism is the total set of chemical processes that keep you alive — building tissues, breaking down fuel, maintaining gradients, generating heat. A few precise terms:
- Basal metabolic rate (BMR) — the energy your body uses at complete rest, just to stay alive: running the heart, the brain, the kidneys, and — recall from the cellular energy page — the constant pumping of the sodium-potassium pump in every cell, which alone accounts for a large chunk of resting energy use. BMR is the bulk of most people's daily energy expenditure.
- Thermogenesis — heat production. Recall uncoupling from the cellular energy page: mitochondria can deliberately leak their proton gradient as heat instead of making ATP. The amount of this background heat-making is a real, adjustable component of metabolic rate. "Running hot" is literally this.
- Substrate flux — which fuel is being used (glucose vs fat, from the Randle cycle), and whether the body is in storage mode or breakdown mode. Metabolism is not just a speed; it is a direction — building up (anabolic) or breaking down (catabolic).
So "metabolic rate" is really a combination of how much machinery you are running (mitochondria, ion pumps), how much heat you are deliberately wasting (uncoupling), and which direction your fuel is flowing. The hormones on this page are the controls that set all three. The master control is the thyroid.
The thyroid: the body's metabolic thermostat
The thyroid is a butterfly-shaped gland in the neck, and its hormones set the overall tempo of metabolism in nearly every cell. But the thyroid does not act on its own whim — it sits at the bottom of a three-tier control chain with feedback, exactly like a thermostat.
The HPT axis — the control chain
The chain is called the HPT axis (hypothalamus–pituitary–thyroid), and it works top-down:
- The hypothalamus (a control hub at the base of the brain that monitors the body's state) releases TRH (thyrotropin-releasing hormone). TRH is a message to the next tier down.
- The pituitary gland (a small gland just below the hypothalamus, the body's "master gland") responds to TRH by releasing TSH (thyroid-stimulating hormone) into the blood.
- The thyroid gland responds to TSH by producing and releasing its hormones, chiefly T4 (and some T3).
Then comes the crucial part — the feedback loop. The thyroid hormones circulating in the blood travel back up to the hypothalamus and pituitary and tell them to ease off: high thyroid hormone suppresses TRH and TSH. This is negative feedback — the output inhibits its own production, exactly like a thermostat shutting off the heating once the room is warm enough. It keeps thyroid hormone in a stable band.
flowchart TD
HYP[Hypothalamus] -->|TRH| PIT[Pituitary gland]
PIT -->|TSH| THY[Thyroid gland]
THY -->|"T4 (mostly) + some T3"| HORM[Thyroid hormone in blood]
HORM -.->|negative feedback:<br/>'enough — ease off'| PIT
HORM -.->|negative feedback| HYP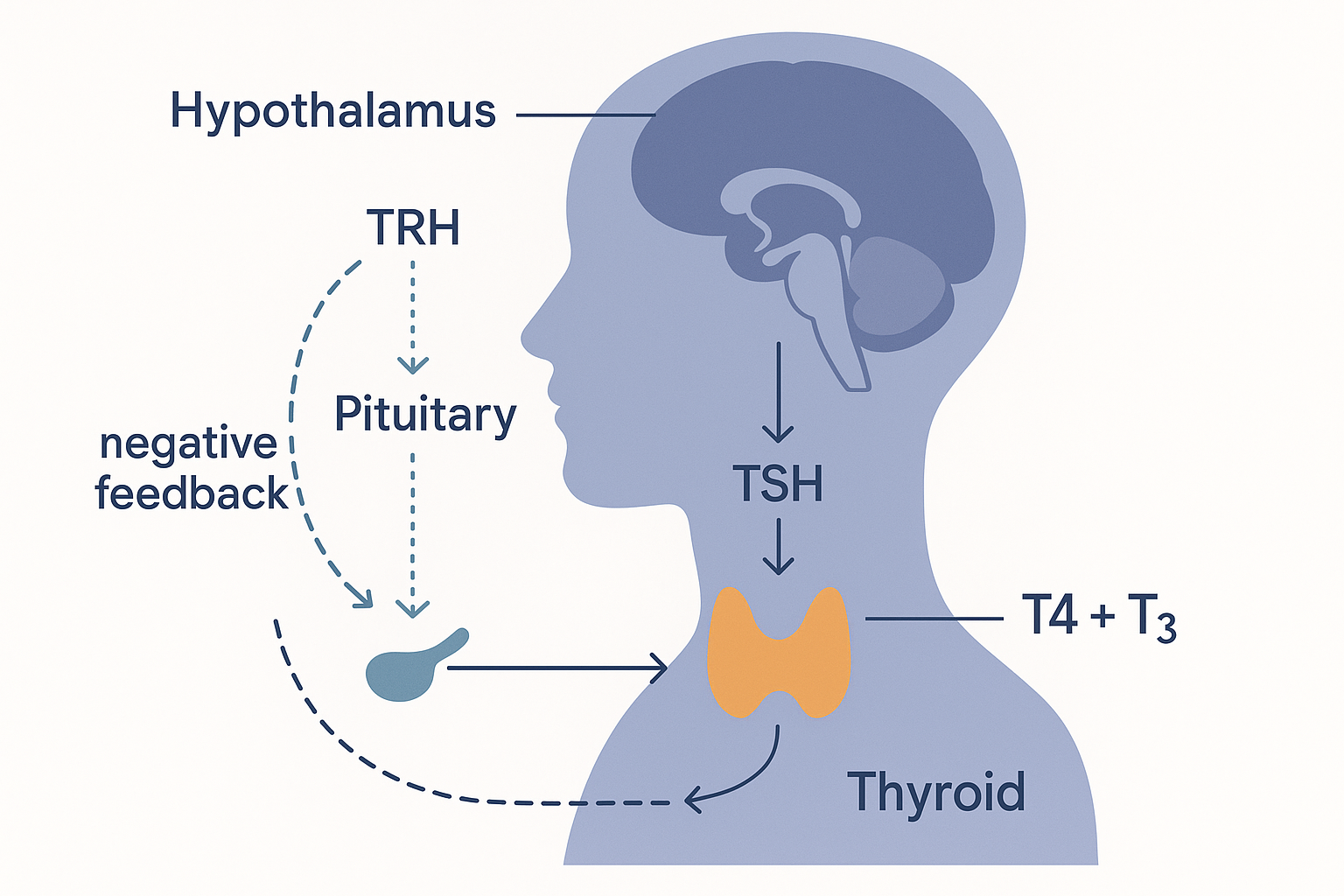 The thyroid thermostat: a top-down chain (hypothalamus → pituitary → thyroid) with negative feedback that holds thyroid hormone in a stable band — and the reason a high TSH signals an under-active thyroid.
The thyroid thermostat: a top-down chain (hypothalamus → pituitary → thyroid) with negative feedback that holds thyroid hormone in a stable band — and the reason a high TSH signals an under-active thyroid.
This feedback loop is also why the standard blood test for thyroid function measures TSH, not thyroid hormone directly. If thyroid hormone is low, the pituitary shouts louder (TSH goes up) trying to stimulate the gland; if thyroid hormone is high, TSH goes down. So a high TSH usually signals an under-active thyroid. (Hold onto a caveat for later: this loop watches T4 levels, but the active hormone is T3 made elsewhere — so the test can read "normal" while the active signal is actually low.)
T4, T3, and T2 — the hormones themselves
The thyroid's hormones are named by how many iodine atoms they carry — and this detail is the key to everything:
- T4 (thyroxine) carries four iodine atoms. It is the main product of the thyroid (~90% of output) — but it is largely a prohormone: a storage/transport form that binds the thyroid receptor only weakly and does little on its own. Think of it as the inactive reserve.
- T3 (triiodothyronine) carries three iodine atoms — one fewer than T4. It is the active hormone, binding the receptor with roughly ten times the affinity of T4. T3 is what actually drives metabolism. Critically, most T3 is not made by the thyroid; it is made by removing one iodine from T4, out in the tissues — especially the liver.
- T2 (diiodothyronine) carries two iodine atoms. Long dismissed as an inert breakdown product, T2 is now thought to have direct effects on mitochondria — speeding them up without going through the slow gene-transcription route that T3 uses. This is still an area of active research and some uncertainty, but it is mechanistically interesting because it would be a faster, more direct metabolic lever than T3.
The pattern to remember: iodine count = activity level, and the body activates thyroid hormone by stripping iodine off, step by step (T4 → T3 → T2).
Activating the hormone — T4 → T3 conversion (and its sabotage)
This step was introduced on the liver page; here is the fuller picture. Converting inactive T4 into active T3 is done by enzymes called deiodinases (they remove iodine — "de-iodinase"). There are three types, and the difference matters:
- DIO1 — mainly in the liver and kidneys; produces T3 for the whole body's circulation. Needs selenium.
- DIO2 — inside specific target tissues (brain, pituitary, muscle, brown fat); makes T3 locally, right where it is needed, for that tissue's own use. This is how some tissues control their own thyroid exposure independent of the blood level.
- DIO3 — the off switch. It converts T4 into reverse T3 (rT3) — an inactive mirror-image isomer that fits the receptor but does nothing, occupying the space like a key that turns no lock. It also inactivates T3 directly.
Under good conditions, the body favours DIO1/DIO2 and makes plenty of active T3. But under stress, illness, fasting, inflammation, or selenium deficiency, the body shifts toward DIO3 and reverse T3 — deliberately applying the metabolic brake. This is an adaptive response: in famine or serious illness, slowing the metabolism conserves energy. The problem is when the trigger is chronic (ongoing psychological stress, chronic inflammation, perpetual dieting) — then the brake stays on indefinitely, producing the picture of low active T3 despite a normal-looking thyroid gland and normal T4.
What inhibits healthy conversion, in a list worth memorising:
- Cortisol (chronic stress) — directly suppresses DIO1 and promotes reverse T3.
- Inflammation (including from gut endotoxin — a link to the gut microbiome page).
- Fasting / chronic calorie restriction — the body reads low intake as famine and down-regulates T3.
- Selenium deficiency — no cofactor, no conversion.
- High oestrogen — raises the binding protein that holds thyroid hormone, lowering the free, usable fraction.
- Liver dysfunction — fewer functioning hepatocytes to do the converting.
How thyroid hormone actually speeds up metabolism
Here is the mechanistic heart of it — what "T3 increases your metabolism" actually means at the molecular level. T3 enters cells and binds thyroid hormone receptors inside the cell nucleus. These receptors are transcription factors — proteins that switch genes on. When T3 binds, it turns up the production of specific proteins, and which proteins it boosts is the whole story:
- More mitochondria — T3 drives mitochondrial biogenesis, the building of new mitochondria. More power stations per cell means more capacity to burn fuel. (Connect to cellular energy: more of the entire electron-transport-chain machinery.)
- More sodium-potassium pumps (Na⁺/K⁺-ATPase) — T3 increases the number of these pumps. Since these pumps are one of the single biggest consumers of ATP at rest, making more of them directly raises baseline energy expenditure. The cell is, in effect, choosing to run more energy-burning pumps.
- More uncoupling proteins — T3 increases proton-leak channels, so more of the proton gradient is dissipated as heat rather than captured as ATP. This is why thyroid hormone makes you warmer and why hypothyroid people feel cold.
flowchart TD
T3[T3 enters cell] --> NUC["Binds thyroid receptor<br/>in the nucleus<br/>(a gene switch)"]
NUC -->|turns genes up| MITO[More mitochondria<br/>= more fuel-burning capacity]
NUC -->|turns genes up| PUMP["More Na+/K+ pumps<br/>= more ATP spent at rest"]
NUC -->|turns genes up| UCP[More uncoupling proteins<br/>= more heat]
MITO --> RATE[↑ Metabolic rate]
PUMP --> RATE
UCP --> HEAT[↑ Body heat]
HEAT --> RATE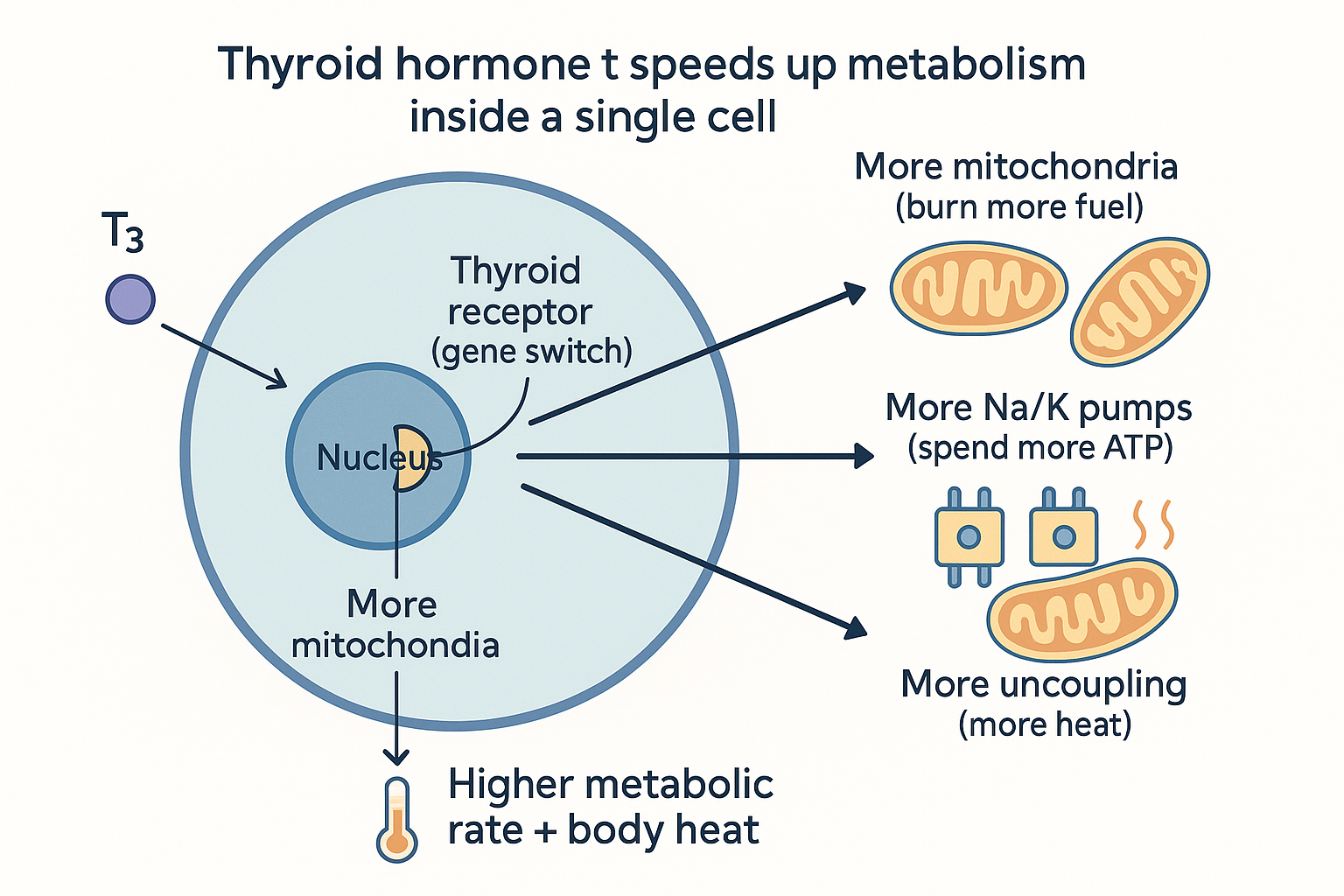 What "T3 speeds up metabolism" means mechanically: T3 is a gene switch that tells the cell to build more fuel-burning machinery and deliberately waste more energy as heat.
What "T3 speeds up metabolism" means mechanically: T3 is a gene switch that tells the cell to build more fuel-burning machinery and deliberately waste more energy as heat.
So "fast metabolism" from thyroid hormone is not magic — it is the cell, instructed by T3, building more machinery and deliberately wasting more energy as heat. A faster metabolism genuinely costs more fuel because the cell is running more pumps and leaking more gradient. This also explains the slow onset: because it works through gene transcription and building new proteins, thyroid effects take days to weeks to manifest, not minutes.
Why the thyroid gets down-regulated
Putting the inhibitors together, the thyroid system is suppressed by:
- Chronic stress (cortisol) — both at the conversion step and centrally.
- Fasting and chronic dieting — the famine signal.
- Systemic inflammation — including low-grade inflammation from gut dysfunction.
- Iodine deficiency — no raw material to build the hormone (the thyroid literally builds T4 around iodine atoms).
- Selenium deficiency — no cofactor to activate it.
- Autoimmune disease (Hashimoto's) — the immune system attacks and destroys the gland itself, the most common cause of hypothyroidism in iodine-replete countries.
- Pituitary dysfunction — if the pituitary fails to send TSH, the thyroid is never told to produce.
Notice how many of these are not problems with the thyroid gland itself — they are problems with its inputs (iodine, TSH), its activation (selenium, liver, cortisol), or its environment (inflammation). This is why thyroid problems are so often really problems elsewhere in the system.
Insulin: the nutrient-partitioning hormone
If thyroid sets the tempo, insulin decides where fuel goes. Released by the pancreas when blood glucose rises (after a meal), insulin is the master "store it" signal. Its message to the body: fuel is plentiful right now — take it out of the blood, use it, and store the surplus. Specifically, insulin:
- Opens the glucose doors — triggers GLUT4 transporters to the surface of muscle and fat cells so glucose can enter (from the cellular energy page).
- Drives storage — tells the liver to store glucose as glycogen and to build fat; tells muscle to build glycogen and protein.
- Suppresses fat release — strongly blocks lipolysis (the breakdown of stored fat). This is critical: while insulin is high, you cannot meaningfully burn body fat, because the release valve is held shut. This is the hormonal logic behind why constant snacking (constant insulin) impedes fat loss.
So insulin is fundamentally anabolic (building/storing) and the antagonist of fat-burning. It is balanced by glucagon (its mirror-image from the liver page) which says "release fuel" when blood sugar falls.
Insulin resistance — when the signal stops working
Insulin resistance is the central dysfunction in much of modern metabolic disease, and it has a precise mechanism. Normally insulin binds its receptor on a cell, which triggers an internal relay (via a protein called IRS-1, insulin receptor substrate 1) that ultimately moves the glucose doors into place. In insulin resistance, this relay is jammed: the insulin binds, but the internal signal fails to get through, so the glucose doors don't open properly. Glucose stays in the blood.
What jams the relay? The main culprits:
- Ectopic fat — fat stored inside muscle and liver cells (where it doesn't belong) produces signalling molecules that block the IRS-1 relay (specifically by causing the wrong kind of chemical tag — serine phosphorylation — on IRS-1, which deactivates it).
- Inflammation — inflammatory signals (including endotoxin/LPS from a leaky gut — the gut microbiome link again) do the same thing to IRS-1.
- Hyperinsulinaemia itself — chronically high insulin (from constant carbohydrate intake) down-regulates its own receptors, so the resistance becomes self-reinforcing: high insulin → resistance → the pancreas makes even more insulin to compensate → more resistance.
The body's response to resistance is to pump out more insulin to force the glucose in — which keeps blood sugar normal for years (so a glucose test looks fine) while insulin levels climb silently. By the time blood sugar finally rises (type 2 diabetes), the problem has been building for a long time. And because high insulin blocks fat-burning, insulin resistance tends to lock in fat storage.
Cortisol and the HPA axis: the stress controller
Cortisol is the body's main long-term stress hormone, a glucocorticoid (the name itself means "glucose + cortex + steroid" — a steroid from the adrenal cortex that raises glucose). Its job is to prepare the body to deal with a challenge by mobilising energy:
- Raises blood glucose — drives the liver's gluconeogenesis (making new glucose, from the liver page), ensuring fuel for the brain and muscles during stress.
- Mobilises fatty acids — releases fat from storage for fuel.
- Suppresses the immune system — useful short-term (this is why synthetic glucocorticoids like prednisone are anti-inflammatory drugs), harmful when chronic.
- Blunts T4 → T3 conversion — the direct link to the thyroid section: cortisol applies the metabolic brake.
- Redistributes fat to the visceral (belly) depot — chronic cortisol promotes the dangerous fat-around-the-organs pattern.
Like the thyroid, cortisol is governed by a three-tier feedback axis — the HPA axis (hypothalamus–pituitary–adrenal):
- The hypothalamus releases CRH (corticotropin-releasing hormone).
- The pituitary responds with ACTH (adrenocorticotropic hormone).
- The adrenal glands (on top of the kidneys) respond by releasing cortisol.
- Negative feedback: cortisol travels back up and suppresses CRH and ACTH — shutting off its own production once the threat passes.
flowchart TD
HYP[Hypothalamus] -->|CRH| PIT[Pituitary gland]
PIT -->|ACTH| ADR[Adrenal glands]
ADR -->|Cortisol| EFF["Raise glucose · mobilise fat ·<br/>suppress immunity ·<br/>blunt T4→T3 · visceral fat"]
EFF -.->|negative feedback| PIT
EFF -.->|negative feedback| HYP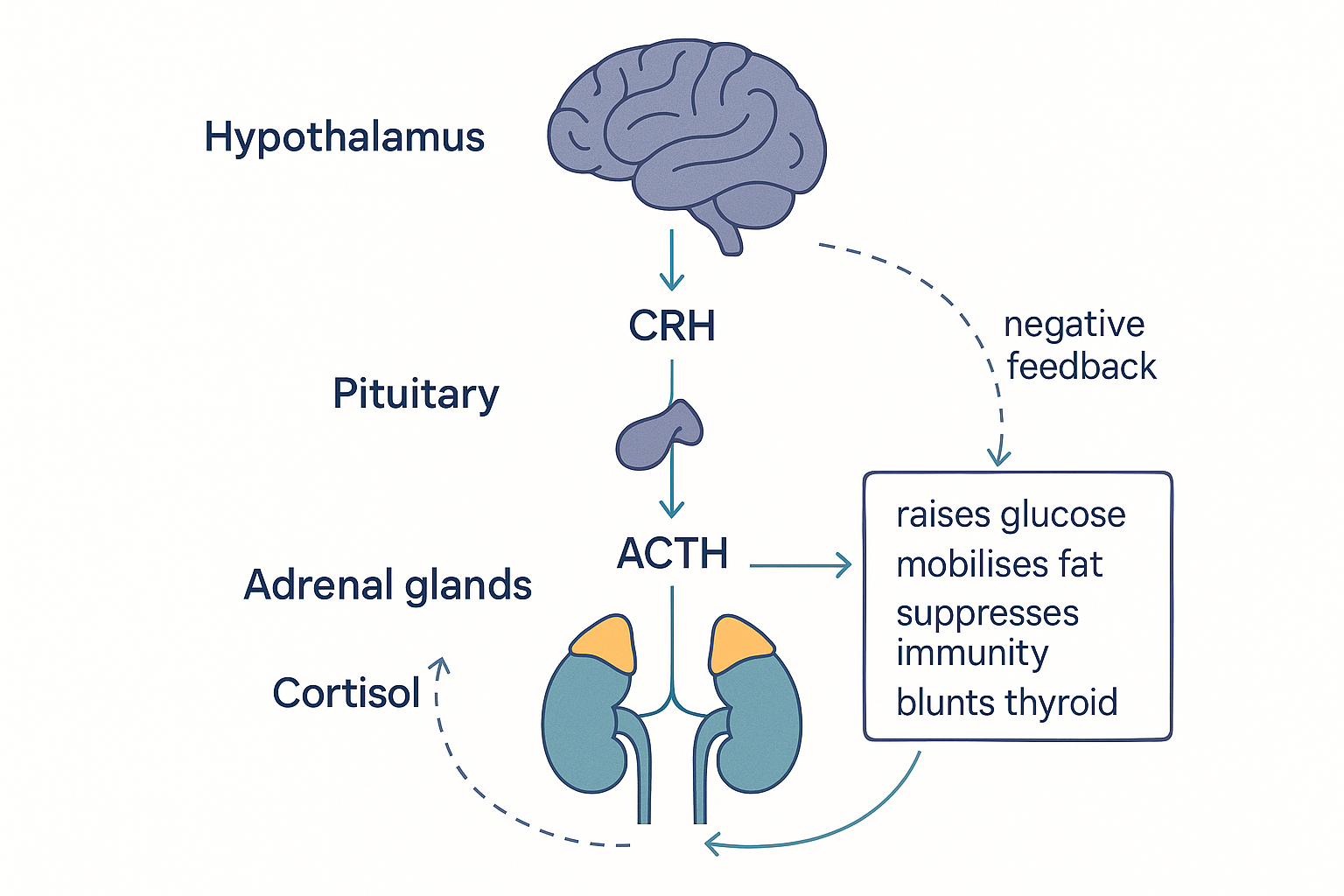 The stress axis, built like the thyroid axis: hypothalamus → pituitary → adrenals, with cortisol feeding back to shut itself off — designed for acute threats, damaging when stuck on.
The stress axis, built like the thyroid axis: hypothalamus → pituitary → adrenals, with cortisol feeding back to shut itself off — designed for acute threats, damaging when stuck on.
The HPA axis is designed for acute stress: spike cortisol, deal with the threat, shut off. The modern problem is chronic activation — ongoing psychological stress keeps the axis switched on, cortisol stays elevated, and the feedback can become dysregulated. Sustained high cortisol is where the cross-system damage happens, as we'll see in the master loop.
Leptin and ghrelin: the hunger and satiety signals
Two more hormones regulate how much you eat, completing the picture:
- Leptin is the satiety ("I'm full / I have enough energy stored") hormone. It is released by fat cells in proportion to how much fat you carry — so it is a signal of long-term energy stores. More body fat → more leptin → the brain (hypothalamus) registers "energy reserves are full" and reduces hunger and permits a normal metabolic rate.
- Ghrelin is the hunger hormone, released mainly by the stomach when it is empty. Ghrelin rises before meals and falls after eating — a short-term "the tank is empty, go eat" signal.
In a healthy system these balance appetite against energy stores. But two failure modes matter:
- Leptin resistance — exactly parallel to insulin resistance. In obesity, leptin is chronically high (lots of fat making lots of leptin), and the hypothalamus stops responding to it. The brain no longer "hears" the satiety signal, so it behaves as if starving — driving hunger and lowering metabolic rate even amid abundant stores. This is a key reason obesity is self-reinforcing rather than self-correcting.
- The dieting trap — when you lose fat, leptin drops (less fat, less leptin), and the body interprets the fall as a famine warning: hunger rises, metabolic rate falls, and thyroid output is trimmed. This is the hormonal basis of the notorious weight-loss plateau and rebound — the body actively defends its fat stores by down-regulating metabolism.
Putting the system together: the master loops
Now the payoff — the engineer's view of how these wire together. The individual hormones are not independent dials; they are a connected control system, and the connections explain why metabolic problems cluster together.
The central vicious loop — chronic stress:
flowchart TD
STRESS[Chronic stress / poor sleep /<br/>inflammation / over-dieting] --> CORT[↑ Cortisol]
CORT -->|blunts DIO1, ↑ reverse T3| LOWT3[↓ Active T3<br/>metabolic brake on]
CORT -->|jams IRS-1 relay| IR[↑ Insulin resistance]
GUT[Leaky gut → endotoxin/LPS] -->|inflammation| IR
GUT -->|inflammation| LOWT3
IR -->|high insulin blocks fat-burning| FATSTORE[↑ Fat storage,<br/>↑ visceral fat]
FATSTORE -->|ectopic + inflammatory fat| IR
FATSTORE -->|chronically high leptin| LEPR[Leptin resistance]
LEPR -->|brain senses 'starving'| LOWMETAB[↓ Metabolic rate,<br/>↑ hunger]
LOWT3 --> LOWMETAB
LOWMETAB -.->|fatigue, more stress| STRESS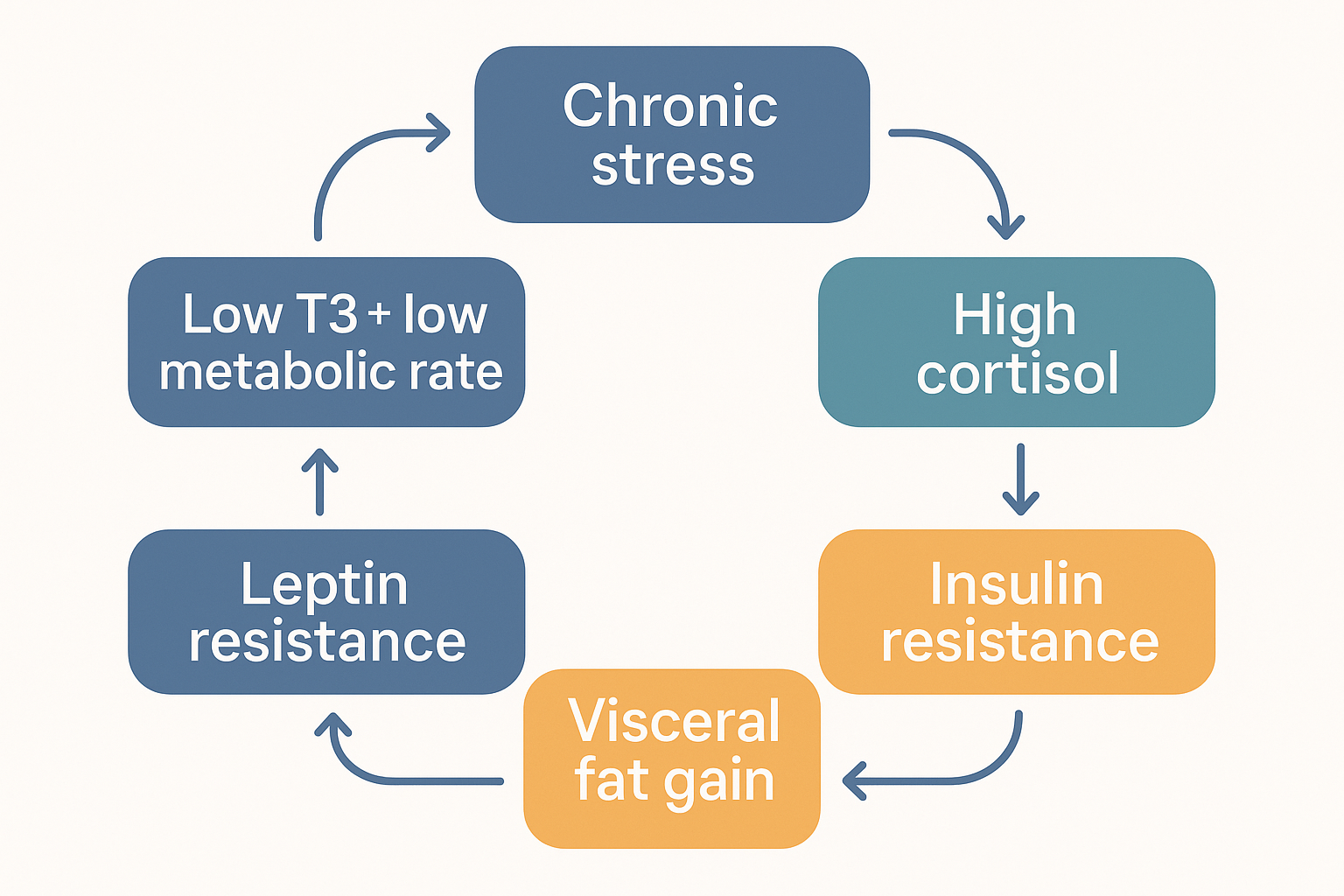 The engineer's view: the major metabolic problems are not separate — they are nodes in one self-reinforcing ring, which is why they cluster together and why breaking the loop at any point helps.
The engineer's view: the major metabolic problems are not separate — they are nodes in one self-reinforcing ring, which is why they cluster together and why breaking the loop at any point helps.
Read it as a system. Chronic stress raises cortisol, which simultaneously suppresses active T3 (slowing the metabolic tempo) and promotes insulin resistance (jamming fuel into storage). Insulin resistance and the resulting visceral fat feed back to worsen insulin resistance (ectopic fat jams IRS-1) and drive leptin resistance (chronically high leptin stops being heard). Both low T3 and leptin resistance lower metabolic rate and raise hunger — which produces fatigue, weight gain, and more stress, closing the loop. Gut dysfunction feeds inflammation into several nodes at once.
This is why the major metabolic problems — fatigue, weight gain, insulin resistance, low thyroid, poor sleep, increased hunger — so rarely appear alone. They are nodes in one connected loop, and pushing any node tends to push the others. It is also why interventions that seem unrelated (improving sleep, lowering stress, healing the gut, gentle refeeding after over-dieting) can break the cycle from multiple directions: they are all targeting the same loop.
The healthy mirror-image: adequate sleep and low chronic stress keep cortisol in check, which permits good T4→T3 conversion (fast tempo) and good insulin sensitivity (fuel used rather than stored), which keeps fat stores moderate and leptin signalling intact (appetite matched to need), which supports a normal metabolic rate and steady energy — a self-reinforcing loop in the good direction.
Putting it all together
- Metabolism is not one number. It is how much machinery you run (mitochondria, ion pumps), how much energy you waste as heat (uncoupling), and which direction fuel flows (store vs burn).
- Thyroid sets the tempo. The HPT axis (hypothalamus → pituitary → thyroid) makes mostly inactive T4; the body activates it to T3 by stripping iodine (mainly in the liver, needing selenium), and T3 raises metabolism by building mitochondria, making more sodium-potassium pumps, and increasing heat-producing uncoupling. Stress, fasting, inflammation, and nutrient deficiency apply the brake (reverse T3).
- Insulin partitions fuel. It is the "store it" signal that opens glucose doors and blocks fat-burning; insulin resistance (jammed IRS-1 relay from ectopic fat and inflammation) is the central modern dysfunction and is self-reinforcing.
- Cortisol controls the stress response via the HPA axis (hypothalamus → pituitary → adrenal); useful acutely, damaging chronically — it suppresses thyroid and drives insulin resistance.
- Leptin and ghrelin regulate appetite; leptin resistance (parallel to insulin resistance) makes the brain sense starvation amid plenty, and dieting-induced leptin drop defends fat stores.
- It is all one loop. Chronic stress, low T3, insulin resistance, leptin resistance, and visceral fat are wired together — which is why metabolic problems cluster, and why fixing sleep, stress, gut, and fuel timing can break the cycle from several angles at once.
The single idea to carry away: metabolism is a feedback-regulated control system, not a furnace with a dial. The hormones sense the body's state and adjust storage, tempo, heat, and hunger to match — and the failure mode is not a broken dial but a loop stuck in the wrong configuration.
Related Compounds & Deep Dives
Thyroid hormone & conversion
- T4 and T3 — the prohormone and the active hormone at the centre of metabolic tempo.
- Selenium — essential cofactor for the deiodinase enzymes that convert T4 to active T3.
- Tyrosine — the amino acid backbone the thyroid builds its hormones on (T4/T3 are iodinated tyrosines).
- Iron and zinc — cofactors required for thyroid hormone production and conversion; deficiency blunts thyroid function.
Stress / HPA axis
- Cortisol — the stress hormone that blunts T4→T3 conversion and drives insulin resistance and visceral fat.
- Pregnenolone, progesterone — steroid hormones upstream of and interacting with the cortisol pathway.
Insulin & fuel partitioning
- Insulin — the "store it" signal; central to nutrient partitioning and insulin resistance.
- Glucose — the fuel whose blood level insulin and the liver jointly regulate.
- Intermittent fasting and exercise — primary levers for restoring insulin sensitivity (and, used to excess, potential triggers of the famine/low-T3 response).
Sex hormones (interacting with the loop)
- Estrogen — raises thyroid-binding protein and lowers free thyroid hormone; interacts with the metabolic loop.
- Testosterone — influences insulin sensitivity and body composition.
Related foundations
- Liver — where T4 is converted to active T3 and where cortisol drives gluconeogenesis.
- Cellular Energy — the mitochondria, sodium-potassium pumps, and uncoupling that thyroid hormone scales up to raise metabolic rate.
- Gut Microbiome (forthcoming) — the source of the endotoxin/inflammation that feeds insulin resistance and low T3.