The Neuroscience of Cognition: Dopamine, Focus, and the Stimulant Landscape
How the brain produces focus, motivation, and wakefulness at the molecular level — and what Adderall, Ritalin, modafinil, caffeine, and the precursor-and-cofactor stacks actually do, mechanism by mechanism, so you can tell which lever each one pulls.
Why this page exists
This is the page that makes the cognitive-enhancement world coherent. People take tyrosine, caffeine, modafinil, Adderall, racetams, and magnesium all "for focus," but these act through completely different mechanisms — some build the raw material, some block its breakdown, some force its release, some remove a brake, some tune the receiver. Lumping them together as "stimulants" hides the whole story. By the end of this page you should be able to look at any focus compound and say which specific step in the dopamine system it touches — and therefore what it will and won't do, and why some are far more addictive or harsh than others.
We build the dopamine system from raw material to receptor, then go through each drug class showing exactly where it intervenes. Along the way we connect to the autonomic nervous system (noradrenaline is shared between brain and body) and the cellular energy page (the brain's enormous glucose appetite).
The cast of chemicals
A few neurotransmitters dominate cognition. Recall from the ANS page that a neurotransmitter is the chemical a neuron releases to signal the next cell, across a tiny gap called the synapse.
- Dopamine — the central character: the chemistry of motivation, focus, and "this is worth pursuing." The star of this page.
- Noradrenaline (norepinephrine) — closely related to dopamine (it is literally made from dopamine), governing alertness and arousal. The same molecule the sympathetic nervous system uses in the body.
- Acetylcholine — attention, learning, and memory encoding (and, from the ANS page, the parasympathetic transmitter).
- Glutamate — the brain's main excitatory signal, central to learning (its NMDA receptor is the subject of the magnesium deep dive).
- Adenosine — an inhibitory "you're getting tired" signal that builds up through the day. The target of caffeine.
Dopamine and noradrenaline are together called catecholamines, and most cognitive stimulants act on them. So we start there.
What dopamine actually is — and isn't
Dopamine is popularly called the "pleasure chemical." This is misleading. Dopamine is better understood as the chemistry of motivation and salience — it signals "this is important, pursue it, this is worth effort" rather than the pleasure of the reward itself. It is the wanting, not the liking. This distinction matters enormously: drugs that flood dopamine do not create happiness, they create compulsive wanting — which is exactly why they are addictive without being satisfying.
Crucially, dopamine is not one system but three distinct pathways, each running from a small source region to a different destination, doing a different job:
- The nigrostriatal pathway (from the substantia nigra to the striatum) controls movement. This is the pathway that dies in Parkinson's disease — which is why Parkinson's is treated with the dopamine precursor L-DOPA.
- The mesocortical pathway (from the ventral tegmental area, or VTA, to the prefrontal cortex) controls focus, working memory, and executive function. This is the pathway that matters most for cognition and the one that cognitive enhancers target.
- The mesolimbic pathway (from the VTA to the limbic system, the nucleus accumbens) is the reward and motivation circuit. This is the pathway drugs of abuse hijack — flooding it produces craving and reinforcement.
So when a compound "raises dopamine," the key question is always which pathway, and how selectively. A drug that selectively boosts the mesocortical (focus) pathway is a clean cognitive enhancer; one that floods the mesolimbic (reward) pathway is an addictive drug. Most real compounds do some of both, and the ratio is what determines their character.
flowchart LR
SN[Substantia nigra] -->|nigrostriatal| STR["Striatum<br/>= MOVEMENT<br/>(dies in Parkinson's)"]
VTA[VTA] -->|mesocortical| PFC["Prefrontal cortex<br/>= FOCUS / executive<br/>(target of cognitive enhancers)"]
VTA -->|mesolimbic| NAC["Nucleus accumbens<br/>= REWARD / craving<br/>(hijacked by drugs of abuse)"]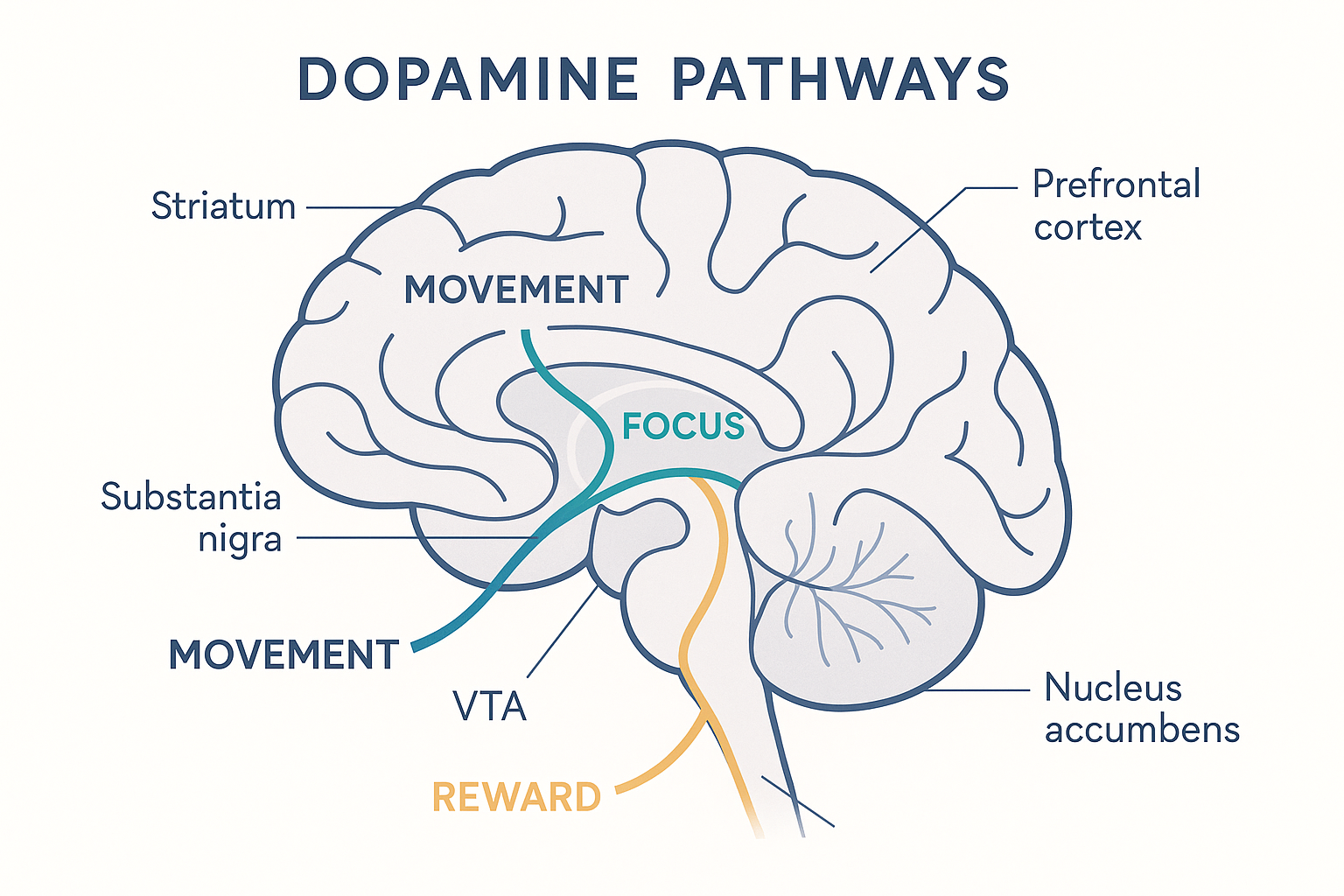 Dopamine is not one system but three separate pathways — which is why "raises dopamine" is meaningless until you ask which pathway: movement, focus, or reward.
Dopamine is not one system but three separate pathways — which is why "raises dopamine" is meaningless until you ask which pathway: movement, focus, or reward.
In the prefrontal cortex, dopamine is co-released with noradrenaline, and the two together tune the quality of focus — a point that becomes central when we reach the inverted-U.
The synthesis chain: building dopamine from food
Dopamine is built step by step from an amino acid you eat, and each step needs a specific enzyme and cofactor. This chain is worth knowing precisely, because half the "focus supplements" are simply feeding or unblocking one of these steps.
- Phenylalanine → Tyrosine. The dietary amino acid phenylalanine is converted to tyrosine (you also get tyrosine directly from protein).
- Tyrosine → L-DOPA. This is the rate-limiting step — the bottleneck that controls the whole pathway's speed. The enzyme is tyrosine hydroxylase, and it requires the cofactors BH4 (tetrahydrobiopterin) and iron. Being rate-limiting means this step sets the ceiling: the pathway can only run as fast as tyrosine hydroxylase allows.
- L-DOPA → Dopamine. The enzyme DOPA decarboxylase converts L-DOPA to dopamine, requiring vitamin B6 (in its active form, P5P).
- Dopamine → Noradrenaline. In noradrenergic neurons, dopamine is further converted to noradrenaline by an enzyme requiring copper and vitamin C.
flowchart LR
PHE[Phenylalanine] --> TYR[Tyrosine]
TYR -->|"tyrosine hydroxylase<br/>★ rate-limiting ★<br/>needs iron + BH4"| DOPA[L-DOPA]
DOPA -->|"needs vitamin B6 (P5P)"| DA[Dopamine]
DA -->|"needs copper + vitamin C"| NA[Noradrenaline]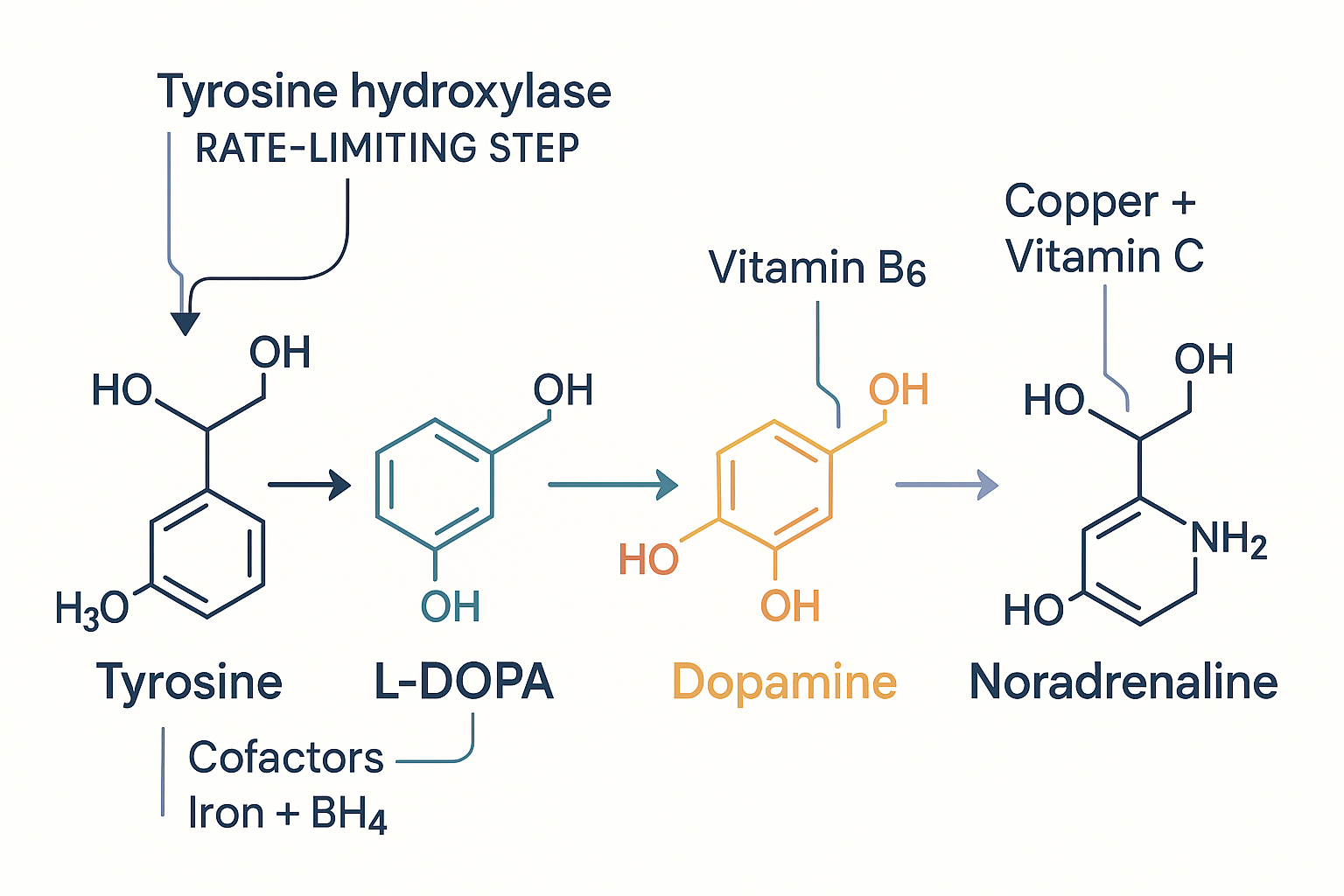 Dopamine is built from the amino acid tyrosine in a cofactor-dependent chain — the first step is the rate-limiting bottleneck, which is why supplementing tyrosine only helps when that step is being run hard.
Dopamine is built from the amino acid tyrosine in a cofactor-dependent chain — the first step is the rate-limiting bottleneck, which is why supplementing tyrosine only helps when that step is being run hard.
This chain explains L-tyrosine supplementation precisely. Tyrosine is the raw material entering the rate-limiting step. Supplementing it raises catecholamine production — but only when production is actually substrate-limited, which happens in states of high demand or depletion: acute stress, sleep deprivation, cold, intense cognitive load. Under normal rested conditions you already have plenty of tyrosine, so adding more does little (the bottleneck is the enzyme, not the raw material). This is why tyrosine "works" for some people in demanding situations and does nothing for others at baseline — it can only help when the tank is being drained faster than diet refills it. It also explains why the cofactors matter: without iron, B6, and copper, the chain cannot run regardless of how much tyrosine you supply. The biohacker stack of "tyrosine + iron + B-vitamins" is really just supplying the substrate and the tools for this one pathway.
The synapse: how a dopamine signal starts and stops
To understand the drugs, you must understand the moment of signalling itself. Here is what happens at a single dopamine synapse:
- The upstream (presynaptic) neuron stores dopamine in tiny packets called vesicles.
- When it fires, it releases dopamine from those vesicles into the synapse — the gap between neurons.
- The dopamine drifts across and binds receptors on the downstream (postsynaptic) neuron, passing the signal on.
- The signal must then be stopped — and how it is stopped is the key to everything. The main "off switch" is reuptake: a protein on the presynaptic neuron called the dopamine transporter (DAT) physically pumps the dopamine back out of the synapse and into the neuron that released it, to be recycled. (Noradrenaline has its own version, the noradrenaline transporter, NET.)
- A backup disposal route is the enzyme MAO (monoamine oxidase), which chemically breaks dopamine down.
So a dopamine signal's strength and duration depend on how much is released, how long it lingers before reuptake, and how fast it is broken down. Every catecholamine stimulant works by tampering with one of these steps — and which step it targets defines the drug.
flowchart LR
VES[Vesicles store dopamine] -->|neuron fires| REL[Dopamine released into synapse]
REL --> REC[Binds receptors on<br/>postsynaptic neuron = signal]
REC --> DAT["DAT pumps dopamine back<br/>(reuptake = the off switch)"]
DAT --> VES
REL -.->|backup disposal| MAO[MAO breaks it down]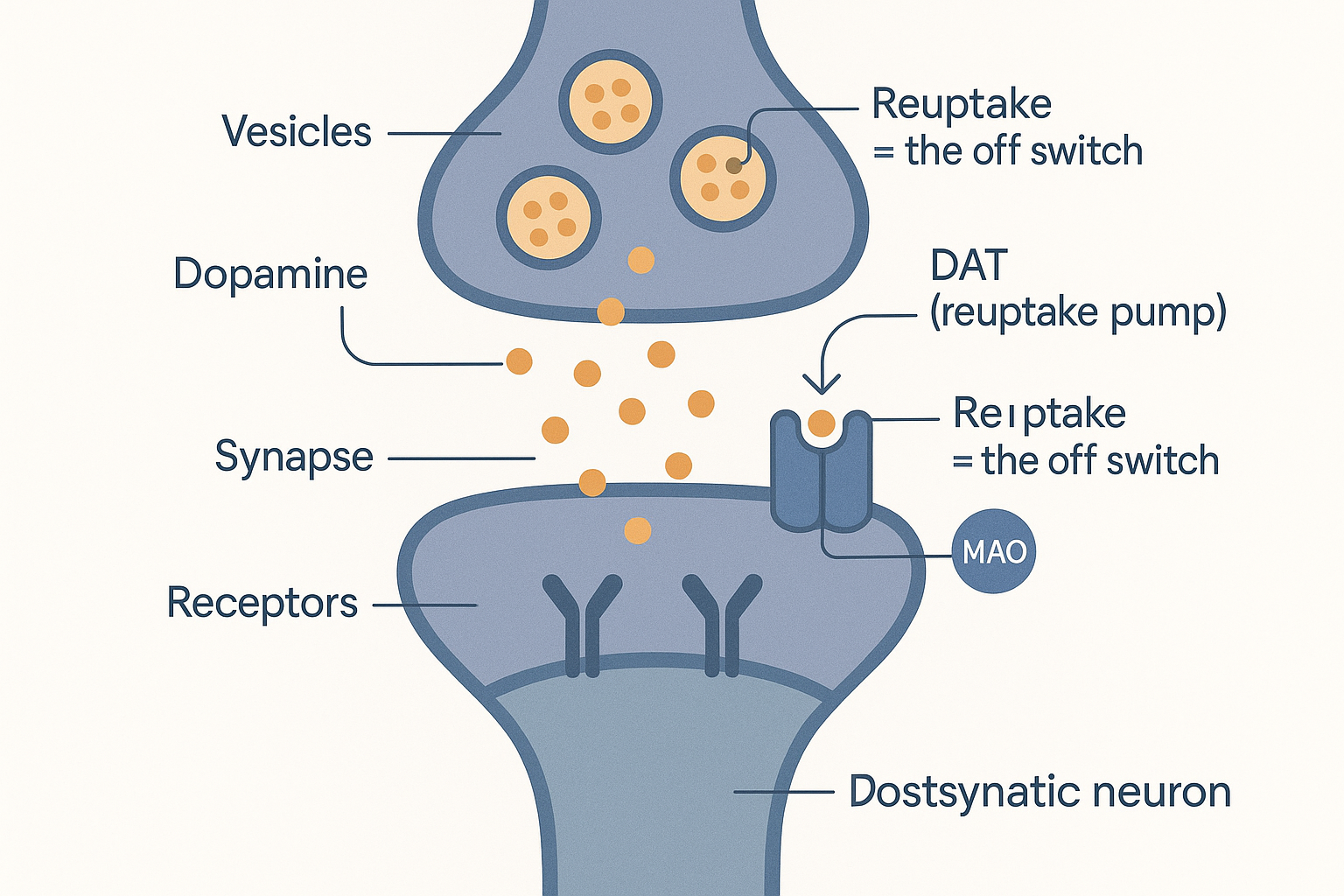 The moment of signalling: dopamine crosses the synapse to the receptors, and is cleared by the DAT reuptake pump — the off switch that nearly every stimulant tampers with.
The moment of signalling: dopamine crosses the synapse to the receptors, and is cleared by the DAT reuptake pump — the off switch that nearly every stimulant tampers with.
The stimulants, mechanism by mechanism
Now the payoff. Four very different things are all called "stimulants." Here is exactly what each does.
Reuptake inhibition — the gentlest lever (methylphenidate / Ritalin)
A reuptake inhibitor blocks the DAT (and NET) — it jams the "off switch." Dopamine that the neuron releases naturally then lingers longer in the synapse, prolonging and amplifying the signal. The critical feature: a pure reuptake inhibitor adds no new dopamine — it only extends what the brain was already releasing. So it amplifies the brain's own signalling pattern rather than overriding it.
Methylphenidate (Ritalin) is essentially a pure DAT/NET reuptake inhibitor. Because it needs dopamine to already be present to have any effect, its action tracks the brain's natural activity — making it more moderate than amphetamine. It is also relatively concentrated in its effect on the prefrontal cortex (the focus pathway): the PFC has low DAT density and depends heavily on reuptake for clearance, so blocking that clearance has an outsized, fairly targeted effect there at low doses. This is the relatively clean "focus" profile.
Reuptake inhibition + forced release — the sledgehammer (amphetamine / Adderall)
Amphetamine (Adderall, dextroamphetamine) does everything methylphenidate does and more — three mechanisms stacked:
- It blocks reuptake (like methylphenidate).
- It reverses the transporters — it makes the DAT run backwards, actively pumping dopamine out of the neuron into the synapse. This is called efflux, and it is the key difference: amphetamine doesn't just preserve released dopamine, it forces dopamine out regardless of whether the neuron was firing.
- It also mildly inhibits MAO, slowing dopamine breakdown.
The result is a much larger dopamine surge that overrides the brain's natural pattern rather than amplifying it. Because it floods the mesolimbic reward pathway as well as the focus pathway — and does so independent of task — it carries a substantially higher abuse potential and a harsher comedown. It is the difference between turning up the volume on the existing signal (methylphenidate) and forcing the speakers to blast regardless of input (amphetamine).
Wakefulness without the flood (modafinil)
Modafinil (and its longer-lasting form armodafinil) is different in character. Its dopamine effect is comparatively weak DAT inhibition — much gentler than methylphenidate — but it adds a distinct wakefulness-promoting action through the brain's orexin/hypocretin and histamine systems (the circuits that keep you awake). The result is strong alertness and wakefulness without the classical dopamine flood, which gives it a much lower reinforcement/abuse potential and a smoother, less euphoric profile. It does not cause the efflux that amphetamine does. This is why it is favoured as a "clean" wakefulness agent rather than a euphoric stimulant. (See the Modafinil & Armodafinil deep dive.)
Removing the brake — a completely different mechanism (caffeine)
Caffeine is not a catecholamine drug at all — it works by an entirely separate route. Throughout the day, the inhibitory molecule adenosine builds up in the brain (a by-product of energy use — recall ATP from the cellular energy page; adenosine is its broken-down remnant). Adenosine binds its receptors and progressively dampens neural activity — this is a major part of why you feel tired as the day goes on. Caffeine is an adenosine receptor antagonist: it blocks those receptors, so adenosine can't apply its brake. By removing the inhibition, caffeine indirectly lets dopamine and noradrenaline signalling proceed more freely — it doesn't add stimulation so much as remove a brake.
This also explains caffeine tolerance: block the adenosine receptors regularly and the brain compensates by making more of them, so you need more caffeine for the same effect, and abrupt withdrawal leaves you with excess adenosine receptors and a crushing tiredness until they downregulate.
The comparison
| Compound | Core mechanism | Adds new dopamine? | Character | Abuse potential |
|---|---|---|---|---|
| Caffeine | Blocks adenosine receptors (removes a brake) | No | Mild alertness, wears with tolerance | Low |
| Modafinil | Weak DAT block + orexin/histamine wakefulness | Barely | Clean wakefulness, not euphoric | Low |
| Methylphenidate | Blocks reuptake (DAT/NET) | No — amplifies own signal | Targeted focus, moderate | Moderate |
| Amphetamine | Reuptake block + forced efflux + MAO inhibition | Yes — forces release | Powerful, overrides natural pattern | High |
The single most useful distinction: does it amplify the brain's own dopamine signalling (caffeine, modafinil, methylphenidate) or override it with forced release (amphetamine)? That line predicts most of the difference in how clean, how powerful, and how addictive each one is.
The inverted-U: why more is not better
Here is the concept that ties the whole stimulant story together and explains why over-dosing focus drugs backfires. The prefrontal cortex's performance does not rise steadily with catecholamine levels. It follows an inverted-U curve: too little dopamine and noradrenaline and the PFC is sluggish and unfocused; the right amount and it performs optimally; too much and performance collapses again — into distraction, anxiety, racing thoughts, and impaired working memory.
flowchart LR
LOW["Too LOW<br/>(fatigue, boredom,<br/>ADHD, under-stimulated)"] --> OPT["OPTIMAL<br/>(calm, focused,<br/>good working memory)"]
OPT --> HIGH["Too HIGH<br/>(stress, anxiety,<br/>racing, scattered)"]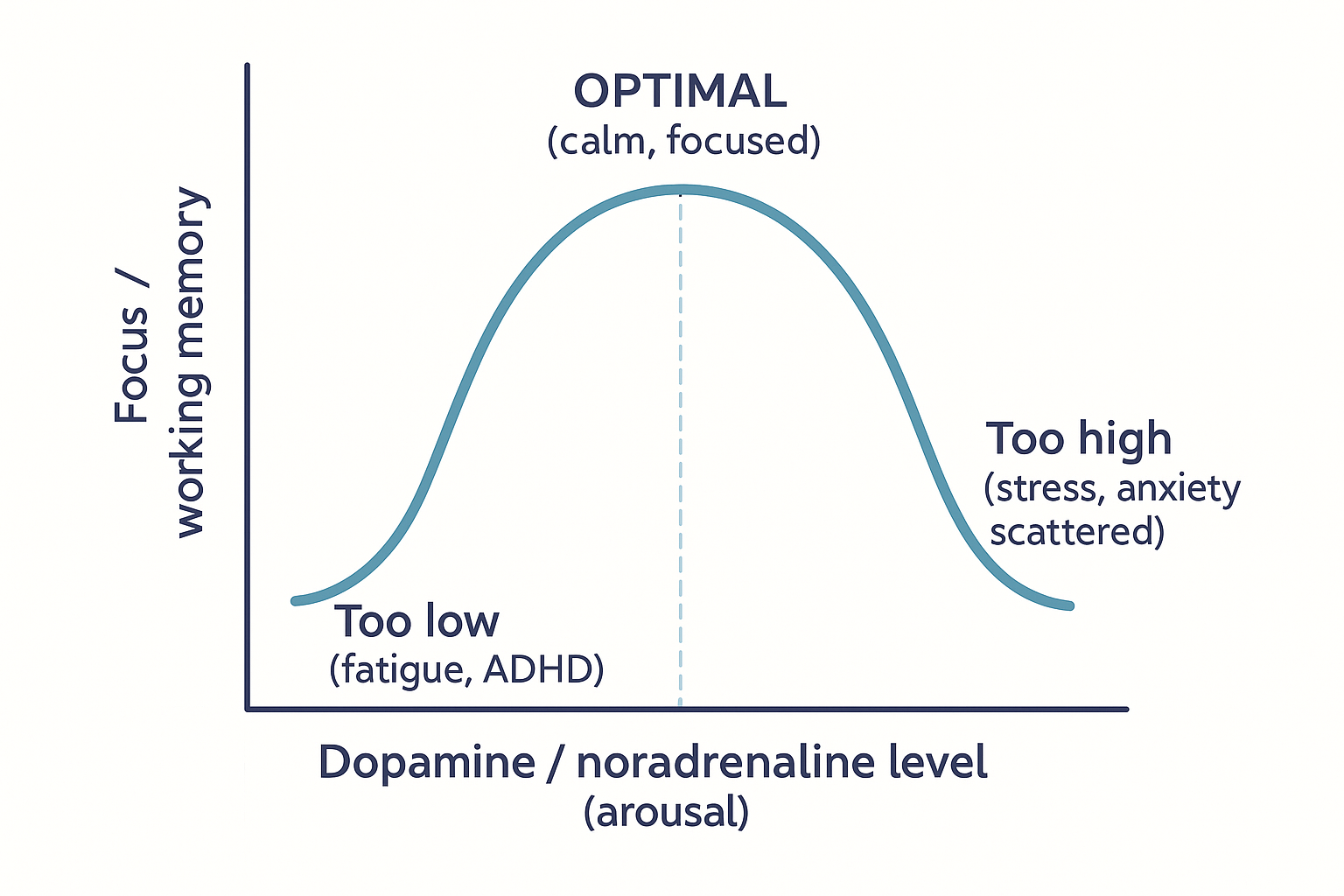 The inverted-U: focus peaks at a moderate catecholamine level. Both too little and too much impair the prefrontal cortex — which is why over-dosing stimulants, and acute stress, both wreck thinking.
The inverted-U: focus peaks at a moderate catecholamine level. Both too little and too much impair the prefrontal cortex — which is why over-dosing stimulants, and acute stress, both wreck thinking.
This single curve explains a remarkable amount:
- Why stimulants help focus at the right dose but wreck it at too high a dose — you can push yourself off the top of the curve down the far side into a wired, scattered, anxious state. More is not better; there is a peak.
- Why stress and anxiety impair thinking even though cortisol and adrenaline are "stimulating." Acute stress floods the PFC with catecholamines, pushing it past the optimum and off the right-hand side of the curve — which is why you "can't think straight" when panicked, blank in an exam, or lose words in an argument. High arousal is not high performance; it is over-shooting the peak. (This connects directly to the ANS and metabolism pages: the sympathetic/cortisol stress response degrades exactly the executive function the PFC is responsible for.)
- Why the goal is the sweet spot, not the maximum. The aim of a cognitive enhancer is to move you to the top of the curve — which means someone under-stimulated (genuine ADHD, fatigue) benefits, while someone already near the peak gets pushed over it and does worse. This is why the same dose helps one person focus and makes another jittery and useless.
Individual differences in baseline catecholamine tone (partly genetic) determine where you sit on this curve to begin with — which is much of why stimulant responses vary so wildly between people.
The brain's fuel: why cognition is metabolically expensive
A thread from the cellular energy page closes an important loop here. The brain is astonishingly energy-hungry: it is about 2% of body weight but consumes roughly 20% of your total glucose. And under normal conditions it runs almost exclusively on glucose — it largely cannot burn fat, because the long-chain fatty acids that fuel muscle do not cross the blood-brain barrier.
The consequences for cognition:
- Stable glucose supply matters for sustained mental work. Hard, prolonged cognition genuinely draws down brain fuel; unstable blood sugar (crashes) produces real cognitive symptoms — fog, irritability, poor focus.
- The one backup fuel is ketones. During prolonged fasting or a ketogenic diet, the liver makes ketone bodies (from the cellular energy page), which can cross the blood-brain barrier and partly fuel neurons. This is why the brain doesn't fail during fasting — but the transition period (before ketone production ramps up) is often when people feel foggy.
- Creatine buffers brain energy too. The phosphocreatine system (cellular energy page) operates in neurons as well as muscle, which is the mechanistic basis for creatine's emerging cognitive benefits — especially under sleep deprivation and high demand, when brain energy is stressed. The popular biohacker claim that "you need good metabolism and glucose for the brain" is, at root, this: cognition is expensive and the brain is a fuel-fussy organ.
Tuning the receiver: magnesium and the NMDA signal
Beyond the chemicals and their fuel, there is the question of signal quality — and here magnesium plays a role covered in depth elsewhere. Briefly: the NMDA receptor (a glutamate receptor central to learning) sits with a magnesium ion plugging its channel, and that magnesium block acts as a signal-to-noise filter — it ensures the receptor only fires for strong, meaningful, coincident signals and ignores random background noise. Adequate magnesium keeps this filter crisp, supporting clean learning and reducing the mental "noise" of anxiety. The full mechanism — and why a specific form crosses into the brain — is in the Magnesium deep dive.
A note on acetylcholine, the other major cognitive transmitter: where dopamine governs motivation and focus, acetylcholine governs attention and the encoding of memory. Its precursors (choline, citicoline) and the nicotinic receptors that nicotine stimulates are the basis of a second family of cognitive compounds aimed at attention and learning rather than drive.
Putting it all together
- Dopamine is motivation/salience ("worth pursuing"), not pleasure — and it runs in three separate pathways: movement (nigrostriatal), focus/executive (mesocortical → PFC), and reward (mesolimbic). Which pathway a drug hits determines whether it's a clean enhancer or an addictive one.
- Dopamine is built in a cofactor-dependent chain (tyrosine → L-DOPA → dopamine → noradrenaline; rate-limited at tyrosine hydroxylase, needing iron, BH4, B6, copper). Supplementing tyrosine helps only when production is substrate-limited (stress, sleep loss).
- Signalling is terminated by reuptake (DAT/NET) and breakdown (MAO) — and every catecholamine stimulant tampers with one of these steps.
- The four "stimulants" are mechanistically distinct: caffeine removes the adenosine brake; modafinil promotes wakefulness with minimal dopamine effect; methylphenidate blocks reuptake to amplify the brain's own signal; amphetamine additionally forces dopamine release (efflux), overriding the natural pattern — which is why it is the most powerful and the most addictive.
- The inverted-U means PFC performance peaks at a moderate catecholamine level — too little or too much both impair, which is why over-dosing stimulants backfires and why stress (over-shooting the peak) makes you unable to think.
- The brain runs almost entirely on glucose (~20% of the body's supply), can fall back on ketones, and is buffered by creatine — cognition is metabolically expensive.
- Magnesium tunes the NMDA signal-to-noise filter, and acetylcholine governs attention and memory encoding — two further levers beyond the dopamine system.
The unifying idea: "focus" is not one thing you can simply turn up. It is the output of a built-from-food dopamine system, terminated by reuptake, tuned to a sweet-spot level in the prefrontal cortex, and powered by a glucose-hungry brain. Each cognitive compound pulls exactly one of these levers — and knowing which lever tells you what it will do, how clean it will be, and where its limits and dangers lie.
Related Compounds & Deep Dives
Dopamine precursors & cofactors
- L-tyrosine — the substrate entering the rate-limiting step; helps when catecholamine production is depleted.
- L-DOPA — the direct dopamine precursor (one step from dopamine).
- P5P (vitamin B6), iron, copper — the cofactors the synthesis chain depends on.
- Dopamine — the neurotransmitter itself.
Stimulants (by mechanism)
- Caffeine — adenosine receptor antagonist (removes the brake).
- Modafinil / armodafinil — wakefulness via orexin/histamine with weak DAT effect (see the deep dive).
- Methylphenidate — reuptake inhibitor; amplifies the brain's own dopamine.
- Adderall / dextroamphetamine — reuptake block plus forced efflux; the most powerful and most addictive.
- Phenylpiracetam — a stimulating racetam with catecholaminergic effects.
- Selegiline — MAO-B inhibitor; slows dopamine breakdown (the disposal route).
- Bromantane — upregulates dopamine synthesis (a different lever; see the deep dive).
Calming the over-shoot / tuning
- L-theanine — smooths caffeine's edge and pulls back from the over-aroused side of the inverted-U.
- Magnesium — NMDA signal-to-noise filter (see the deep dive).
Acetylcholine / attention & memory
- Acetylcholine, choline, citicoline — the attention-and-memory transmitter and its precursors.
- Nicotine — nicotinic receptor agonist for attention (mechanism, not endorsement).
Brain energy
- Creatine — phosphocreatine buffer for neurons under high demand.
- Glucose — the brain's primary fuel.
Related foundations
- Cellular Energy — the brain's glucose appetite, ketones, and the creatine buffer.
- Autonomic Nervous System — noradrenaline shared between brain and body; the stress response that over-shoots the PFC inverted-U.
- Systemic Metabolism — the cortisol/stress system that degrades executive function.