The Gut Microbiome: The Ecosystem Within
The trillions of microbes living in your gut are not passengers — they are a metabolically active organ. What lives there, what "good" and "bad" bacteria actually do, and how a disturbed gut quietly drives inflammation, insulin resistance, low thyroid, and hormone disruption throughout the whole body.
Why this page exists
The previous foundations pages kept pointing here. The liver page noted that the portal vein delivers gut-derived material — including bacterial toxins — straight to the liver. The systemic metabolism page repeatedly blamed "inflammation, including endotoxin from a leaky gut" for insulin resistance and suppressed thyroid conversion. This page is where those references get explained from the ground up.
The central claim, which sounds dramatic but is well-supported, is this: the state of your gut microbiome is one of the most important upstream regulators of whole-body health — not because of vague "gut health" wellness reasons, but through a specific, traceable mechanism in which bacterial toxins leak into the bloodstream and trigger inflammation that degrades metabolism, liver function, and hormone balance. Understanding that mechanism makes an enormous amount of biohacking discourse — endotoxin, leaky gut, probiotics, the fibre debate, "gut healing" — finally coherent.
We build it in order: first what is actually down there, then what the residents do, then the barrier that contains them, then what happens when that barrier fails.
What is actually in the gut
Your gut — principally the colon (large intestine), where the digestion page left off — is home to roughly 38 trillion microorganisms, a number comparable to the count of your own human cells. Collectively this community is called the microbiome (the full set of microbes and their genes) or the gut flora. A few framing facts:
- It is mostly bacteria, but not only bacteria. Alongside bacteria live archaea (a separate ancient class of single-celled organisms, some of which produce methane), fungi (notably Candida, a yeast — the collective fungal population is called the mycobiome), and an enormous population of viruses that mostly infect the bacteria (the phageome). It is a full ecosystem, not a monoculture.
- The bacteria are grouped into a few dominant phyla. A phylum is a broad branch of the bacterial family tree. Two phyla make up roughly 90% of gut bacteria: the Firmicutes and the Bacteroidetes. The ratio between them shifts with diet and is one crude marker of gut state. Smaller but important phyla include the Proteobacteria (which contains many of the troublemakers) and Actinobacteria (which contains the beneficial Bifidobacterium).
- One structural distinction matters more than any other: gram-negative vs gram-positive. Bacteria come in two wall types, named after a staining test. Gram-negative bacteria have an extra outer membrane, and that outer membrane is built partly from a molecule called lipopolysaccharide (LPS) — the endotoxin that is the villain of this entire page. Gram-positive bacteria lack that outer membrane and so do not carry LPS. Keep this distinction in mind: gram-negative = carries endotoxin.
The microbiome is best thought of as a metabolic organ — collectively it carries more genes than your own genome, and it performs chemistry your own cells cannot, from fermenting fibre to manufacturing vitamins.
"Good" vs "bad" bacteria — what that actually means
People talk about good and bad gut bacteria as if it were a morality tale. Mechanistically, the distinction is concrete: it is about what each microbe produces and whether it protects or degrades the gut barrier.
Beneficial bacteria — the ones you want — include genera like Bifidobacterium, Lactobacillus, Akkermansia muciniphila, and Faecalibacterium prausnitzii. What makes them "good" is what they do:
- They produce short-chain fatty acids (SCFAs) — especially butyrate — by fermenting fibre. SCFAs are the gut's most important beneficial output (covered in detail below): they feed the gut lining, tighten the barrier, and calm inflammation.
- They manufacture vitamins — several B vitamins and vitamin K are synthesised by gut bacteria.
- They protect the mucosal barrier. Akkermansia, for instance, lives in the mucus layer and stimulates the gut to maintain it. A healthy barrier keeps the bacteria out of the bloodstream.
- They crowd out the troublemakers. A dense, healthy population of beneficial bacteria physically and chemically occupies the space, leaving no room and no food for harmful species. This is called colonisation resistance — a healthy ecosystem defends itself by being full.
Harmful or opportunistic bacteria are not always "bad" — many are normal residents that only cause trouble when they overgrow. When the balance tips toward them, they produce a very different output:
- LPS (endotoxin) — from the gram-negative species (many Proteobacteria), the inflammatory molecule.
- Hydrogen sulphide and other irritant gases.
- Secondary bile acids — some of which are pro-inflammatory or carcinogenic in excess.
- Zonulin triggers — signals that actively loosen the gut barrier (below), helping the toxins escape.
This unhealthy tipped-over state has a name: dysbiosis (dys- = bad, disordered; the opposite of the healthy balanced state, eubiosis). Dysbiosis is not just "fewer good bacteria" — it is an active shift toward an inflammatory, barrier-degrading community, often involving overgrowth of Proteobacteria, certain Clostridia, or the Candida yeast.
flowchart LR
subgraph EU["EUBIOSIS (healthy balance)"]
G["Bifidobacterium · Lactobacillus<br/>Akkermansia · F. prausnitzii"] --> GOUT["Produce SCFAs + vitamins<br/>protect barrier<br/>colonisation resistance"]
end
subgraph DYS["DYSBIOSIS (tipped over)"]
B["Overgrown Proteobacteria ·<br/>Clostridia · Candida"] --> BOUT["Produce LPS endotoxin · gases ·<br/>secondary bile acids ·<br/>loosen the barrier"]
end
GOUT --> HEALTH[Barrier intact, low inflammation]
BOUT --> SICK[Barrier leaks, inflammation rises]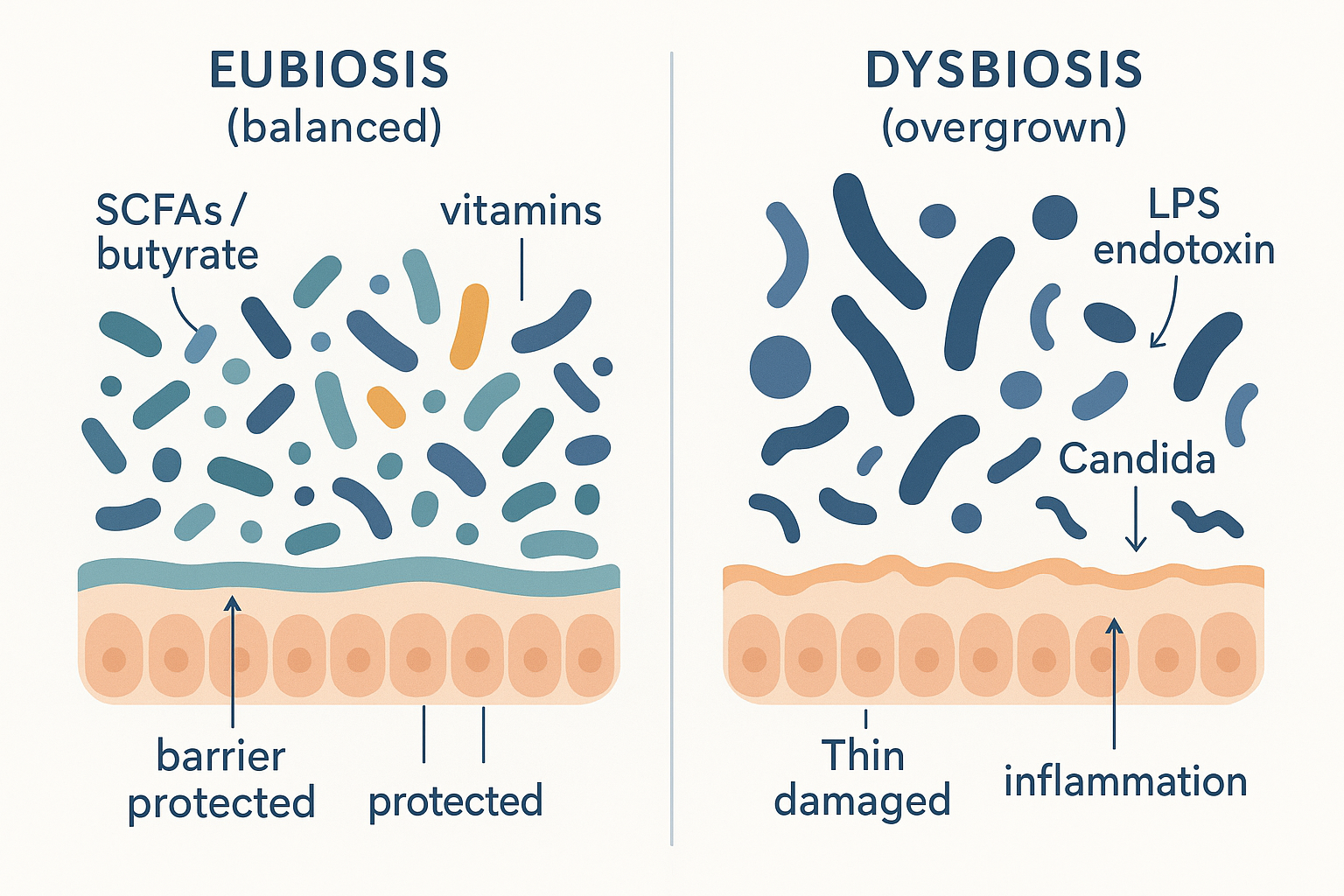 Eubiosis vs dysbiosis: the same space, two very different communities — one diverse and barrier-protecting, the other overgrown and endotoxin-producing.
Eubiosis vs dysbiosis: the same space, two very different communities — one diverse and barrier-protecting, the other overgrown and endotoxin-producing.
The gut barrier: the wall between "outside" and "inside"
Here is a fact that reframes everything: the inside of your gut tube is, topologically, still the outside world. The food and the trillions of bacteria in your gut lumen are separated from your actual bloodstream and tissues by a remarkably thin wall. Keeping that wall intact is the single most important defensive job in the gut, because on the other side is your portal vein — a direct highway to the liver and body.
The barrier has three layers, from the lumen inward:
- The mucus layer. A coat of gel (mucus) lines the gut wall, physically keeping most bacteria at arm's length from the actual cells. Beneficial bacteria like Akkermansia help maintain this layer; it is the first line of defence.
- The epithelial cell layer — one cell thick. This is the astonishing part: the entire barrier between the bacterial soup of your colon and your bloodstream is a single layer of cells (enterocytes/colonocytes). One cell thick.
- The tight junctions. The cells in that single layer are stitched together by protein seals called tight junctions, built from proteins named occludin and claudin. These seals close the gaps between cells, so that anything entering the body must pass through a cell (which can regulate what it admits) rather than slipping between cells unchecked.
A key regulator of these seals is a protein called zonulin: when zonulin is released, it loosens the tight junctions, opening the gaps. Some gut bacteria and certain foods trigger zonulin release — which is the molecular basis of the next concept.
flowchart TD
LUMEN["GUT LUMEN<br/>(bacteria + food = still 'outside')"] --> MUCUS[Mucus layer<br/>keeps bacteria at arm's length]
MUCUS --> CELLS["Epithelial cells — ONE cell thick<br/>sealed by tight junctions<br/>(occludin, claudin)"]
CELLS --> BLOOD["Portal vein → liver → body<br/>('inside')"]
ZON[Zonulin released] -. loosens junctions .-> CELLS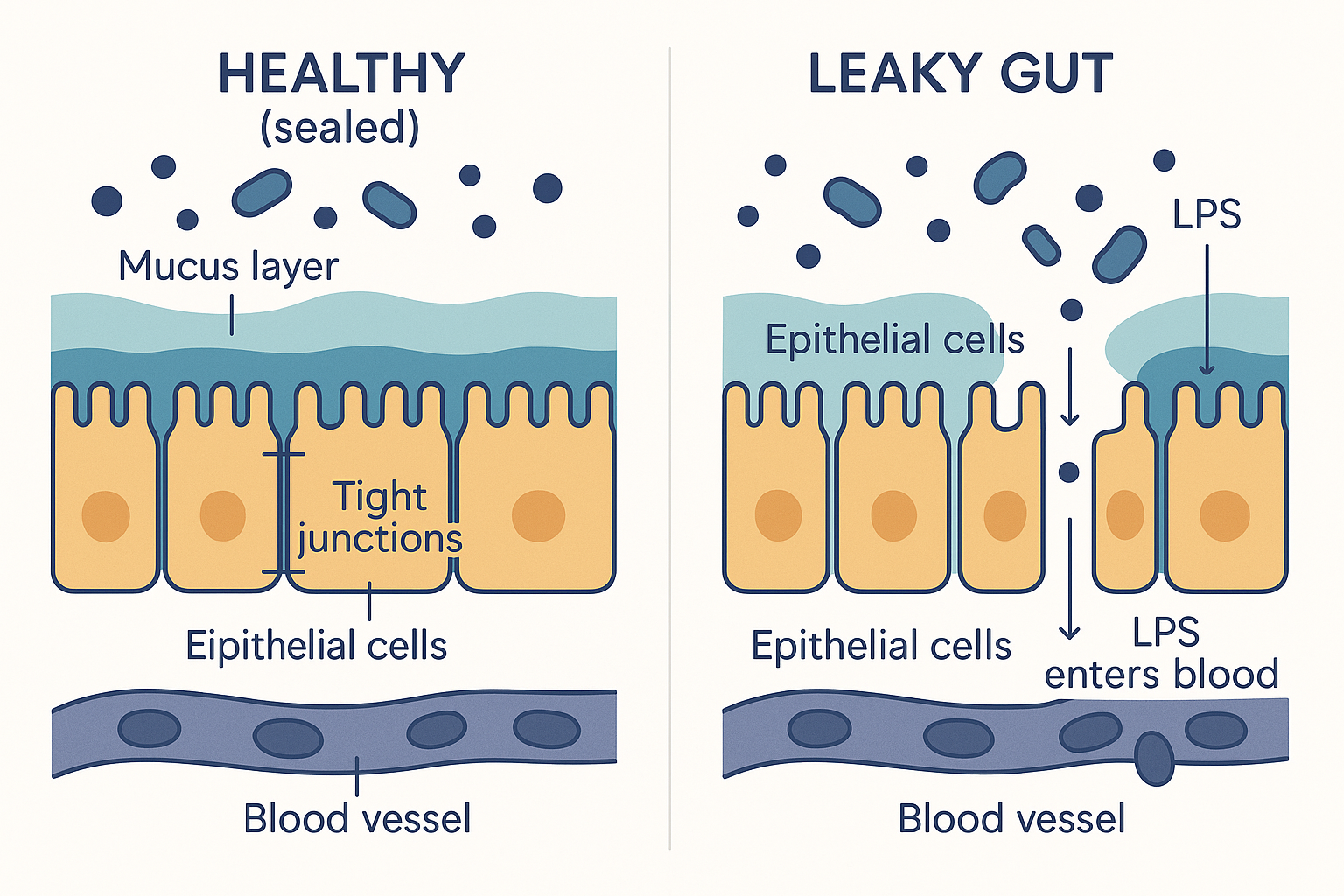 The one-cell-thick wall: when the tight junctions hold, bacteria and endotoxin stay in the lumen; when they loosen (leaky gut), LPS slips through into the bloodstream.
The one-cell-thick wall: when the tight junctions hold, bacteria and endotoxin stay in the lumen; when they loosen (leaky gut), LPS slips through into the bloodstream.
Leaky gut: when the barrier fails
"Leaky gut," more precisely increased intestinal permeability, is the condition where those tight junctions loosen and the one-cell wall becomes porous. Things that should have stayed in the lumen — bacterial fragments, LPS, partially digested food particles — slip between the cells into the tissue and bloodstream beneath.
What disrupts the tight junctions:
- LPS itself — endotoxin triggers zonulin and inflammation that loosen the seals, a vicious circle: leak lets LPS through, LPS widens the leak.
- Dysbiosis — an unhealthy community produces zonulin triggers and fewer barrier-protective SCFAs.
- Alcohol — directly damages the barrier (connecting to the liver page's alcohol story).
- NSAIDs — painkillers like ibuprofen damage the gut lining.
- Gluten — in susceptible people — gliadin (a gluten component) can trigger zonulin release; this is pronounced in coeliac disease and variable in others.
- Chronic stress — high cortisol degrades the barrier (a link to the metabolism page's HPA axis, and a preview of the gut-brain connection).
- A lack of butyrate — because butyrate is the fuel that the barrier cells need to maintain themselves (below).
Once the barrier leaks, the central pathology of this page begins.
Endotoxin (LPS) and metabolic endotoxaemia
LPS (lipopolysaccharide), also called endotoxin, is a fragment of the outer membrane of gram-negative bacteria. It is shed constantly as those bacteria die and divide. Inside the gut lumen, contained behind the barrier, it is harmless — you carry a large reservoir of it at all times with no ill effect. The problem is when it crosses into the bloodstream.
Two routes let it cross:
- Through a leaky barrier — the main route; loosened tight junctions let LPS slip through.
- Piggybacking on dietary fat — LPS is partly fat-soluble, so it can be packaged into chylomicrons (the fat-transport particles from the digestion page) and ferried across with absorbed fat. This is why a very high-fat meal, especially against a dysbiotic gut, transiently raises blood LPS.
When LPS is chronically present in the blood at low levels, the condition is called metabolic endotoxaemia (endotoxin in the blood, at a metabolically disruptive low grade). Here is what it does, step by step:
- LPS is recognised by the immune system as a bacterial invader. Immune cells carry a receptor called TLR4 (toll-like receptor 4) that is specifically tuned to detect LPS. To the body, LPS in the blood means "gram-negative bacteria have breached the wall" — a genuine emergency signal.
- TLR4 activation switches on the master inflammation programme. Binding LPS triggers an internal pathway that activates a control protein called NF-κB (nuclear factor kappa B) — the master switch for inflammatory genes. The cell starts producing inflammatory messengers (cytokines like TNF-α and IL-6).
- The result is chronic, low-grade, system-wide inflammation — not the acute redness of an injury, but a persistent simmer throughout the body. And that simmer is what damages the other systems.
flowchart TD
DYS[Dysbiosis: gram-negative overgrowth] --> LPS[LPS released in gut]
LEAK[Leaky barrier] --> CROSS
FAT[High-fat meal → chylomicrons] --> CROSS
LPS --> CROSS[LPS crosses into blood]
CROSS --> PORTAL["Portal vein → LIVER first<br/>(highest LPS exposure)"]
CROSS --> SYS[Systemic circulation]
PORTAL --> TLR4
SYS --> TLR4["TLR4 receptor on immune cells"]
TLR4 --> NFKB["NF-κB switch → cytokines<br/>(TNF-α, IL-6)"]
NFKB --> INFLAM[Chronic low-grade inflammation]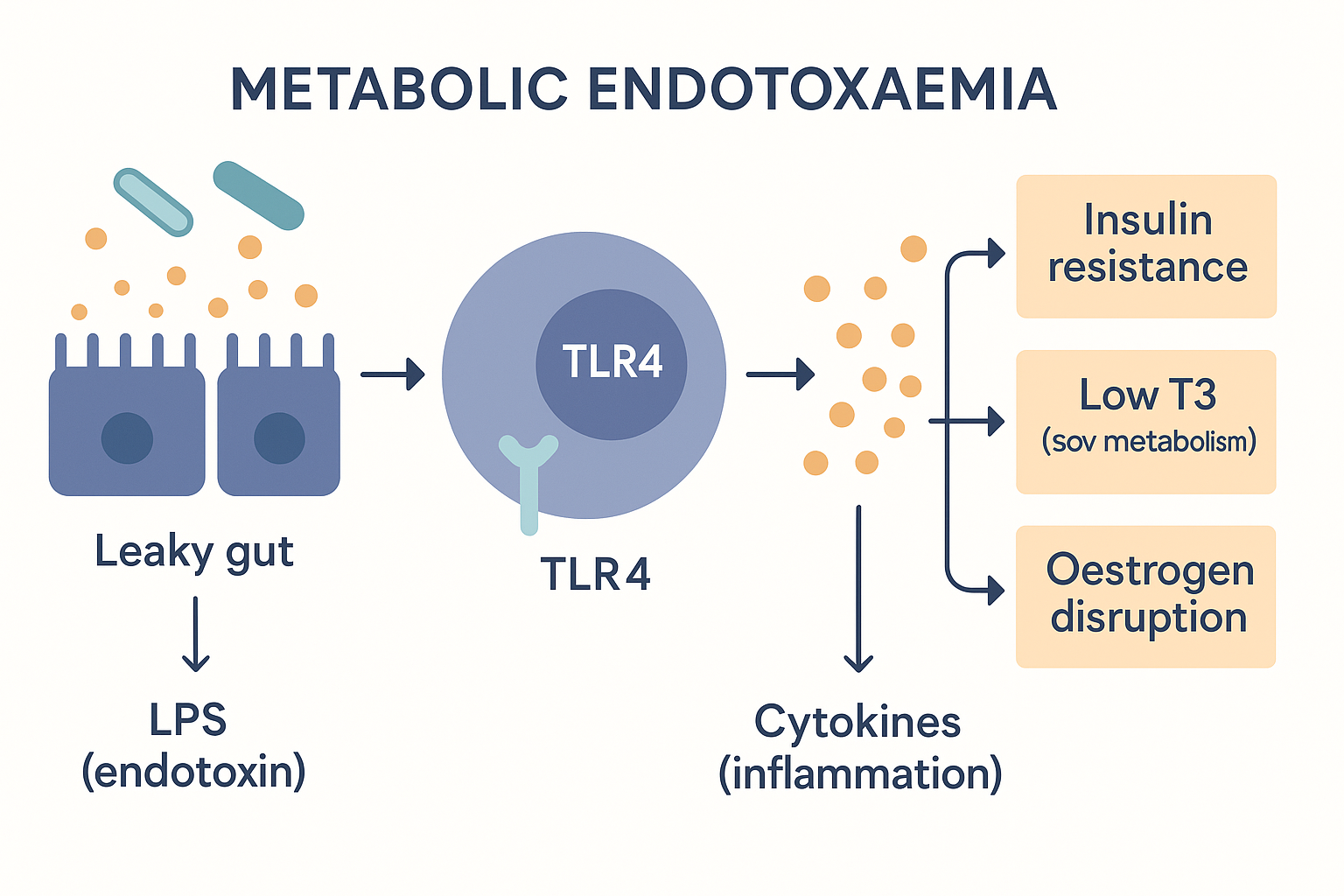 Metabolic endotoxaemia: leaked LPS is read as a bacterial invasion (TLR4 → inflammation), and that inflammation reaches out to cause insulin resistance, low thyroid, and hormone disruption.
Metabolic endotoxaemia: leaked LPS is read as a bacterial invasion (TLR4 → inflammation), and that inflammation reaches out to cause insulin resistance, low thyroid, and hormone disruption.
What metabolic endotoxaemia does to the rest of the body
This is where the gut reaches out and degrades the systems described on the earlier pages. The chronic inflammation from leaked LPS has specific, traceable consequences:
- Insulin resistance. The inflammatory signalling from LPS (via TNF-α) causes the wrong chemical tag on IRS-1 — the exact serine-phosphorylation mechanism described on the metabolism page that jams the insulin relay. So a leaky gut is a direct, upstream cause of insulin resistance. This is one of the most important and underappreciated links in metabolic health.
- Liver inflammation. Recall from the liver page that the portal vein delivers gut blood straight to the liver first. That means the liver receives the highest concentration of any leaked LPS, before the rest of the body. Chronic LPS exposure inflames the liver, contributing to fatty liver progressing to inflammation (steatohepatitis), and impairs the liver's many jobs.
- Suppressed thyroid conversion. Inflammation blunts the liver's T4→T3 conversion and pushes toward reverse T3 (the metabolism page's brake) — so a leaky gut can lower your active thyroid hormone and slow your metabolism, with a normal-looking thyroid gland.
- Mitochondrial dysfunction. Inflammatory signalling impairs mitochondrial function (the cellular energy page's power plants), contributing to fatigue at the cellular level.
- Endocrine and hormone disruption. This is subtle and important. The gut bacteria collectively influence oestrogen levels through what is called the estrobolome — the set of gut bacterial genes that act on hormones. Here is the mechanism: the liver inactivates spent oestrogen by tagging it (glucuronidation, from the liver page) and dumping it into bile for excretion via the gut. But certain gut bacteria produce an enzyme called β-glucuronidase that snips that tag back off, reactivating the oestrogen so it gets reabsorbed instead of excreted (via the enterohepatic circulation). A dysbiotic gut with high β-glucuronidase activity therefore raises the body's oestrogen load. LPS-driven inflammation can also increase aromatase (the enzyme that makes oestrogen) and disturb the broader oestrogen/prolactin balance. This is the mechanistic basis for the claim — common in certain biohacking circles — that gut health influences hormone balance.
Lipid peroxidation — the oxidative damage angle
There is one more consequence worth understanding, because it appears constantly in the Ray Peat–influenced corner of biohacking: lipid peroxidation. Chronic inflammation from endotoxin floods tissues with reactive oxygen species (ROS) — unstable, highly reactive molecules. These ROS attack the polyunsaturated fatty acids (PUFAs) in your cell membranes — the fragile, double-bond-rich fats are especially vulnerable to oxidation. When a PUFA is oxidised, it breaks down into toxic secondary products with names like 4-HNE and MDA (malondialdehyde), which themselves damage proteins and DNA and propagate further damage. So the chain runs: endotoxin → inflammation → ROS → oxidation of membrane PUFAs → toxic breakdown products → more cell damage. This is a major reason the anti-PUFA, anti-endotoxin themes travel together in that community — they are two ends of the same oxidative-damage mechanism.
SCFAs: the beneficial output
Now the good side of the ledger, in detail. The single most valuable thing your beneficial bacteria do is ferment fibre into short-chain fatty acids (SCFAs) — small fat molecules that are anything but waste. Three matter:
- Butyrate — the star. It is the preferred fuel of the colon's own lining cells (colonocytes): they run primarily on butyrate, not glucose. This creates a beautiful dependency — the bacteria feed the very cells that house them. Butyrate also tightens the gut barrier (strengthening tight junctions, the opposite of leaky gut), is potently anti-inflammatory, and even signals to cells via gene regulation. A gut producing plenty of butyrate is largely self-protecting against the entire LPS cascade above. (This is why butyrate is supplemented directly and why resistant starch — fermented into butyrate — is valued.)
- Propionate — absorbed into the portal vein and sent to the liver, where it influences glucose production and cholesterol synthesis.
- Acetate — the most abundant; enters the general circulation and can be used as fuel by muscle and other tissues, and is involved in appetite signalling.
flowchart TD
FIBRE[Dietary fibre] -->|fermented by<br/>beneficial bacteria| SCFA[Short-chain fatty acids]
SCFA --> BUT["Butyrate"]
SCFA --> PROP["Propionate"]
SCFA --> ACE["Acetate"]
BUT --> BUTF["Fuels colon lining ·<br/>tightens barrier ·<br/>anti-inflammatory"]
PROP --> PROPF["To LIVER →<br/>glucose & cholesterol control"]
ACE --> ACEF["To BODY →<br/>fuel + appetite signalling"]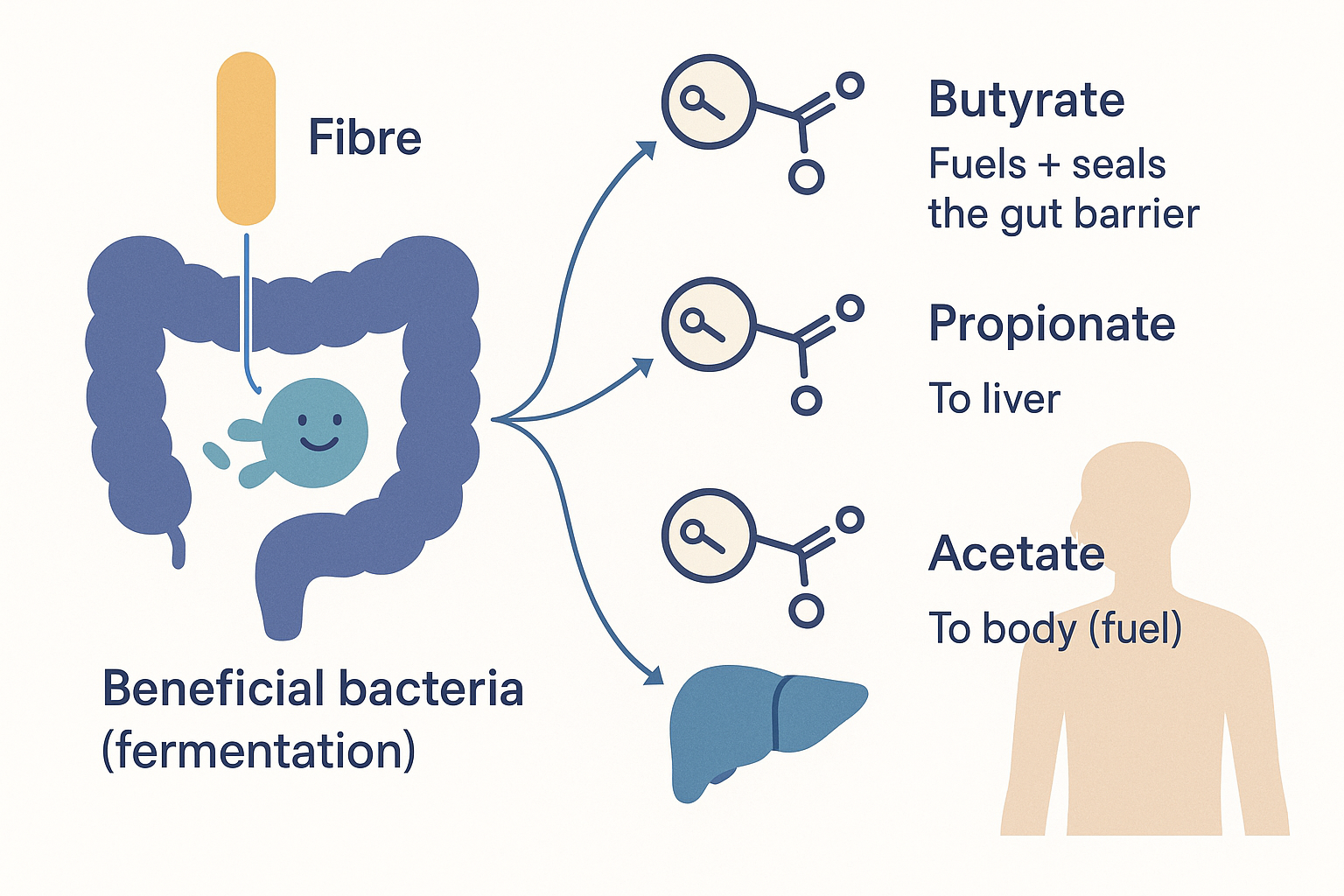 The protective payoff of feeding good bacteria: fibre becomes butyrate (fuels and seals the barrier), propionate (to the liver), and acetate (to the body) — the direct counter to the endotoxin cascade.
The protective payoff of feeding good bacteria: fibre becomes butyrate (fuels and seals the barrier), propionate (to the liver), and acetate (to the body) — the direct counter to the endotoxin cascade.
The takeaway: SCFAs are the mechanism by which "feeding your good bacteria" actually protects you — they fuel and seal the barrier, calm inflammation, and reach the liver and body with beneficial signals. They are the direct antidote to the endotoxin story.
The fibre nuance: not all fibre is equal
Here the page must engage honestly with a genuine controversy. The mainstream view is simple: fibre feeds beneficial bacteria, produces SCFAs, therefore more fibre is better. That is largely true — but there is a well-argued dissenting view (associated with Ray Peat and his followers) that deserves a fair, mechanistic hearing, because the truth is more nuanced than "fibre good."
The nuance is this: fermentable fibre feeds bacteria indiscriminately — both the beneficial and the harmful ones. Highly fermentable soluble fibres (such as inulin, FOS/fructo-oligosaccharides, and pectin) are excellent SCFA substrates, but in a gut that is already dysbiotic or prone to bacterial overgrowth, adding lots of fermentable fibre can also feed the overgrowth — increasing gas, bloating, and potentially endotoxin production. For someone with a healthy gut, fermentable fibre is beneficial; for someone with significant dysbiosis or small-intestinal bacterial overgrowth (SIBO), it can worsen symptoms. This is why blanket "eat more fibre" advice helps some people and makes others distinctly worse.
The Peat-influenced position goes further and distinguishes fermentable fibre from certain non-fermentable, structurally bulky fibres that may act almost as gut antiseptics — physically binding bacteria and endotoxin and helping carry them out, without being fermented into bacterial food. The two classic examples are raw carrot (whose firm, indigestible fibres are thought to bind bacterial toxins and bile in the colon) and cooked white mushroom (similar). The claim is that these reduce endotoxin load without feeding dysbiosis — a different and complementary strategy to "feed the good bacteria with fermentable fibre."
How seriously to take this: the SCFA/butyrate benefits of fermentable fibre are very well established, so this is not a reason to avoid fibre wholesale. But the core insight — that fibre is not monolithic, that fermentation feeds all comers, and that the right fibre strategy depends on the state of your gut — is sound and worth holding. The pragmatic synthesis: a healthy gut tolerates and benefits from fermentable fibre; a dysbiotic gut may need to reduce fermentable load and lean on barrier repair and antiseptic-type fibres first.
What antibiotics do
Antibiotics are the bluntest instrument acting on this ecosystem. A course of broad-spectrum antibiotics does not surgically remove a target pathogen — it carpet-bombs the whole community, killing beneficial and harmful bacteria alike. The consequences flow directly from the concepts above:
- It destroys colonisation resistance. With the dense protective population wiped out, space and food open up — and the organisms that recover fastest are often not the beneficial ones. This permits opportunistic overgrowth: the dangerous Clostridioides difficile (C. diff, which causes severe colitis) and the Candida yeast are classic post-antibiotic overgrowths, because they exploit the cleared field.
- It collapses SCFA production, depriving the colon lining of butyrate and weakening the barrier — which can itself promote leak and inflammation.
- Recovery is slow and often incomplete. The microbiome can take months to recover after a single course, and may never fully return to its prior diversity. Repeated courses compound the loss.
This is not an argument against antibiotics when they are genuinely needed — they are life-saving. It is an argument for using them only when warranted, and for actively rebuilding the ecosystem afterward (with fermented foods, probiotics, and barrier support). The yeast-overgrowth risk is also why antifungals like nystatin sometimes follow antibiotic-driven Candida blooms.
Putting it all together
- The gut is a metabolic organ — ~38 trillion microbes (bacteria, archaea, fungi, viruses), dominated by the Firmicutes and Bacteroidetes phyla. The key structural fact is gram-negative bacteria carry LPS endotoxin; gram-positive do not.
- "Good" bacteria produce SCFAs and vitamins, protect the barrier, and crowd out troublemakers (colonisation resistance); "bad" overgrowth (dysbiosis) produces endotoxin and loosens the barrier.
- The barrier is one cell thick, sealed by tight junctions (occludin, claudin) regulated by zonulin, and coated in mucus. Keeping it intact is the central defensive job — because just behind it is the portal vein to the liver.
- Leaky gut lets LPS cross into the blood (through loosened junctions or piggybacking on dietary fat), producing metabolic endotoxaemia — chronic low-grade inflammation via the TLR4 → NF-κB pathway.
- That inflammation reaches out and degrades the other systems: insulin resistance (jammed IRS-1), liver inflammation (highest LPS exposure via the portal vein), suppressed T4→T3 (slower metabolism), mitochondrial dysfunction, and oestrogen disruption (the estrobolome and β-glucuronidase recycling), plus lipid peroxidation of membrane PUFAs.
- SCFAs (especially butyrate) are the protective antidote — fuelling and sealing the barrier and calming inflammation.
- Fibre is not monolithic — fermentable fibre feeds all bacteria (good and bad), so the right strategy depends on the gut's state; some bulky fibres may act as endotoxin-binding antiseptics instead.
- Antibiotics carpet-bomb the ecosystem, destroying colonisation resistance and inviting C. diff and Candida overgrowth, with slow and often incomplete recovery.
The unifying idea: the gut is the body's hidden upstream control point. A great deal of what looks like a metabolic problem, a thyroid problem, a liver problem, or a hormone problem is downstream of a single root — a disturbed microbiome leaking endotoxin into a body that reads it as a permanent low-grade infection. This is why "fix the gut" is so often the highest-leverage intervention: it sits upstream of nearly everything on the other foundations pages.
Related Compounds & Deep Dives
SCFAs & feeding beneficial bacteria
- Butyrate — the SCFA that fuels and seals the gut barrier; the direct antidote to the endotoxin cascade.
- Resistant starch — fermentable fibre that beneficial bacteria turn into butyrate.
Beneficial bacteria (probiotics)
- Probiotics, VSL#3 — live beneficial strains to rebuild colonisation resistance.
- Lactobacillus, L. reuteri, L. rhamnosus, Akkermansia muciniphila — specific beneficial species; Akkermansia maintains the mucus barrier.
- Saccharomyces boulardii — a beneficial yeast that helps displace pathogens and is used during/after antibiotics.
- Kefir, yogurt, fermented foods — dietary sources of beneficial microbes.
Barrier repair (against leaky gut)
- L-glutamine — primary fuel for enterocytes; supports tight-junction integrity.
- Zinc-L-carnosine, BPC-157 — support healing of the gut lining.
- Larazotide — a zonulin antagonist that directly tightens the junctions.
- Lactoferrin and colostrum — support the barrier and modulate gut bacteria (see the Lactoferrin & Colostrum deep dive).
Managing overgrowth
- Nystatin — antifungal for Candida overgrowth.
- Mastic gum, bismuth — act against specific gut pathogens.
Hormone clearance (the estrobolome link)
- Calcium-D-glucarate — inhibits β-glucuronidase, reducing the reabsorption (recycling) of spent oestrogen.
Protocols & related foundations
- Gut Reset protocol — a practical application of this page.
- Liver — the organ that receives the highest LPS load via the portal vein.
- Systemic Metabolism — how gut-derived inflammation drives insulin resistance and low T3.
- Gut-Brain Axis (forthcoming) — how the microbiome reaches the nervous system.