Vitamin E
Not one molecule but eight. Discovered as a fertility factor, sold as the wrong fraction, and quietly one of the most important things standing between your cell membranes and the slow rust of lipid peroxidation. What vitamin E actually is, how it works at the molecular level, why most supplements are "useless," and how the bioenergetic (Ray Peat) world uses it against inflammation, estrogen, and PUFA damage.
Read alongside
Vitamin E's whole job is to stop lipid peroxidation and the inflammation it drives, so the Inflammation foundations page is the ideal companion. Its mechanism also leans on the membrane and mitochondrial biology of Cellular Energy, and its hormonal effects on the Hormones & Endocrine System page. This page assumes those as background.
Why vitamin E is genuinely mysterious
Most vitamins are a single molecule (vitamin C is ascorbic acid, full stop). Vitamin E is not — it is a family of eight related fat-soluble molecules, and almost every confusing thing about it flows from that fact. The "vitamin E" on a supplement label is usually just one of the eight (and often a synthetic version of that one), which is why people take it and feel nothing, while the bioenergetic crowd insists the full spectrum is transformative. Add a strange origin story (it was discovered as a fertility factor), an unusual mechanism (it works by being sacrificed), and a genuine scientific debate about which fraction matters, and you have a nutrient that even informed people find slippery.
This deep dive takes it apart properly: the chemistry, the history, the eight forms and their wildly different bioavailability, the precise molecular mechanism, and then the Ray Peat framework that treats vitamin E as a frontline tool against inflammation, estrogen, and the damage done by polyunsaturated fats.
What kind of molecule it is
Vitamin E is a fat-soluble (lipophilic) vitamin — meaning, like vitamins A, D, and K (and unlike the water-soluble B and C), it dissolves in fat, not water. From the digestion page, that already tells you a lot: it's absorbed with dietary fat via bile and the lymphatic route, it's stored in the body's fat and membranes rather than excreted quickly, and it must be taken with fat to absorb well.
Structurally, every form of vitamin E shares the same basic design:
- A chromanol ring — the "head," which carries a hydroxyl (–OH) group. This –OH group is the entire functional business end of the molecule — it's the part that does the antioxidant work, as we'll see.
- A lipid tail — a long fatty chain that anchors the molecule inside cell membranes, buried in the fatty layer alongside the very fats it protects.
So picture vitamin E as a molecular harpoon embedded in your cell membranes: tail buried in the fat, antioxidant head poised at the ready. That positioning — inside the membrane, among the vulnerable fats — is the key to everything it does.
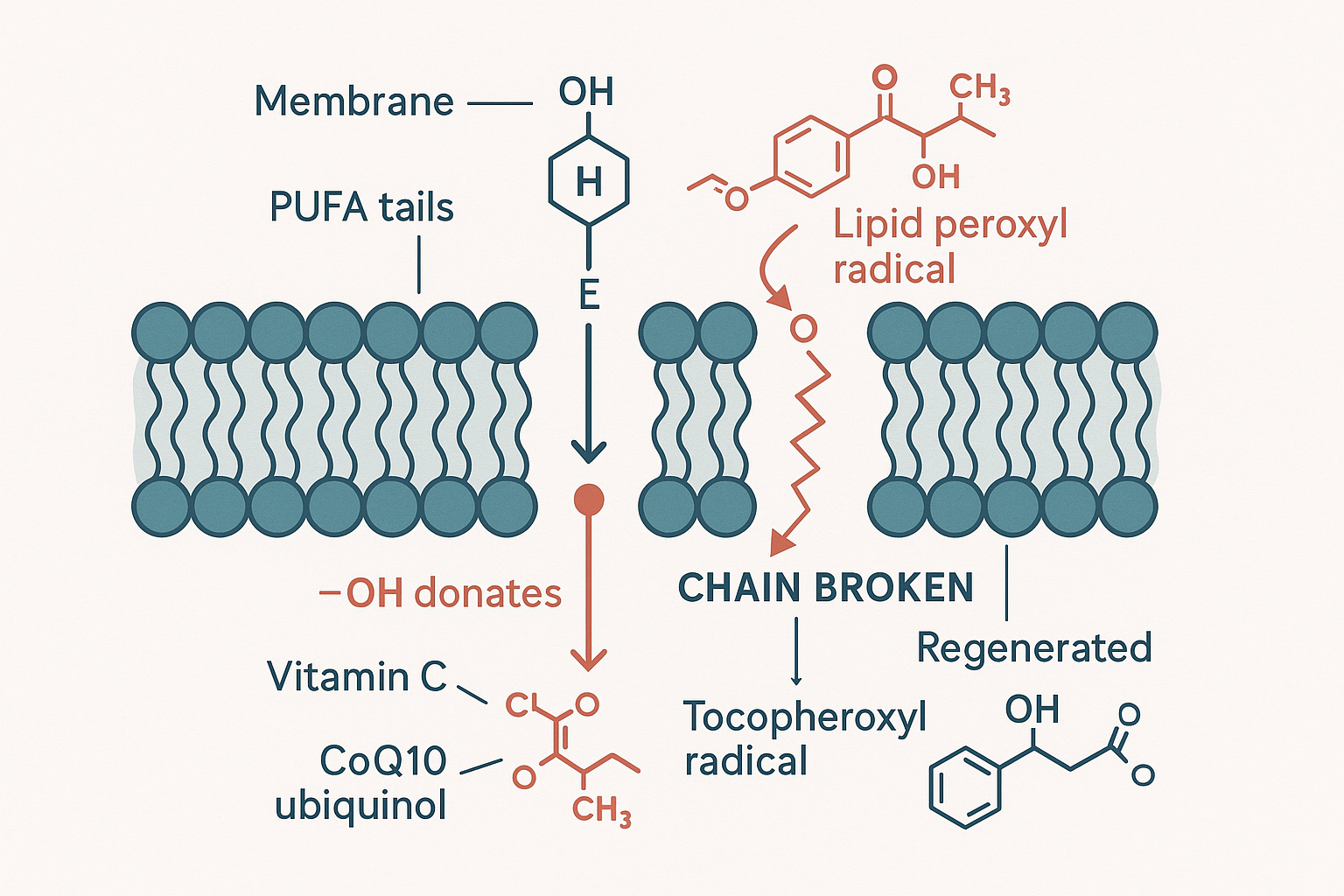 Vitamin E as a molecular harpoon: buried in the membrane among the vulnerable fats, it donates a hydrogen to a lipid peroxyl radical — breaking the chain reaction and taking the bullet — then is regenerated by vitamin C and CoQ10.
Vitamin E as a molecular harpoon: buried in the membrane among the vulnerable fats, it donates a hydrogen to a lipid peroxyl radical — breaking the chain reaction and taking the bullet — then is regenerated by vitamin C and CoQ10.
Is it made in the body? Where it comes from
No — vitamin E is an "essential" nutrient, meaning the body cannot make it and must obtain it from the diet. (This is what "vitamin" means: an essential dietary micronutrient.) And here is a fact the Ray Peat world finds deeply telling: only plants make vitamin E. It is synthesised by photosynthetic organisms — plants, algae — specifically to protect their own polyunsaturated fats from oxidation. When you eat plant fats, you get the vitamin E that came packaged with them to protect them.
The richest natural sources:
- Wheat germ oil — by far the highest (and historically central to this story; see the Vince Gironda note below).
- Seed and vegetable oils — sunflower, safflower, soybean, corn.
- Nuts and seeds — almonds, hazelnuts, sunflower seeds.
- Avocado, spinach, and some other greens (smaller amounts).
Notice the awkward overlap: the foods richest in vitamin E are largely the same seed oils and nuts that the bioenergetic framework tells you to avoid because they are high in PUFA. This is not a coincidence — it's the central irony of vitamin E, and we'll return to it. The plant makes vitamin E to protect its PUFA; the two travel together.
The history: discovered as a fertility factor
Vitamin E's name encodes its origin. In 1922, researchers Herbert Evans and Katharine Scott Bishop at Berkeley found that rats fed an otherwise complete diet became infertile — the females could not carry pregnancies to term. A then-unknown fat-soluble "factor X" in green leaves and wheat germ restored fertility. In 1924 it was named tocopherol, from the Greek tokos (childbirth / offspring) and pherein (to bear) plus -ol (denoting an alcohol, the –OH group) — literally "to bear offspring." Its structure was worked out and it was first synthesised in 1938.
This origin matters for two reasons. First, it permanently tied vitamin E to reproduction and hormones in researchers' minds — a thread the bioenergetic world picks up (vitamin E and steroidogenesis, below). Second, the mid-century enthusiasm — notably the Canadian Shute brothers, who used high-dose vitamin E for heart disease — seeded a tradition of therapeutic, high-dose vitamin E that mainstream medicine later treated with scepticism but that the Ray Peat world inherited and kept alive.
The eight forms — and why this is the whole story
Vitamin E is not one molecule but eight naturally occurring forms (vitamers), split into two families of four:
- Tocopherols — with a saturated (straight, stable) tail. Four types: alpha (α), beta (β), gamma (γ), delta (δ).
- Tocotrienols — with an unsaturated tail (three double bonds). Same four: α, β, γ, δ.
The four members within each family differ only by how many methyl groups decorate the chromanol head — and that small difference changes their chemistry and biology substantially.
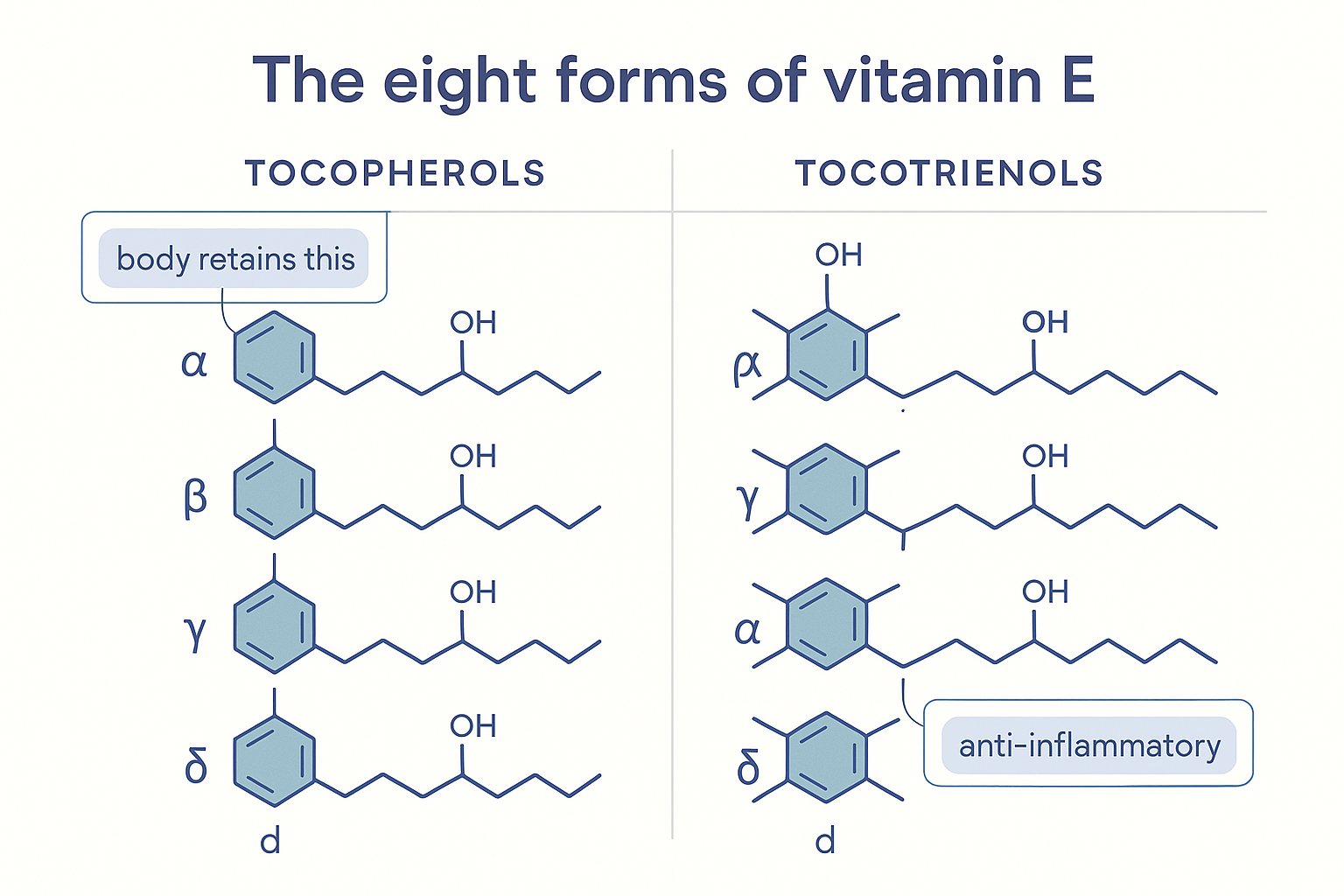 The whole story in one picture: four tocopherols (saturated tail) and four tocotrienols (unsaturated tail), differing only by the methyl groups on the chromanol head — alpha is the form the body retains, gamma the anti-inflammatory fraction.
The whole story in one picture: four tocopherols (saturated tail) and four tocotrienols (unsaturated tail), differing only by the methyl groups on the chromanol head — alpha is the form the body retains, gamma the anti-inflammatory fraction.
flowchart TD
VE["Vitamin E<br/>(8 natural forms)"] --> TOC["TOCOPHEROLS<br/>(saturated tail)"]
VE --> TT["TOCOTRIENOLS<br/>(unsaturated tail)"]
TOC --> A1["α-tocopherol<br/>← the 'official' vitamin E,<br/>body retains this one"]
TOC --> B1["β-tocopherol"]
TOC --> G1["γ-tocopherol<br/>← the anti-inflammatory fraction"]
TOC --> D1["δ-tocopherol"]
TT --> A2["α-tocotrienol"]
TT --> B2["β-tocotrienol"]
TT --> G2["γ-tocotrienol"]
TT --> D2["δ-tocotrienol<br/>(tocotrienols: cholesterol,<br/>hair, neuroprotection)"]Here is the pivotal fact that explains why "vitamin E" is so confusing. Your body strongly prefers one of the eight: alpha-tocopherol. The liver contains a protein called α-tocopherol transfer protein (α-TTP) whose job is to selectively grab α-tocopherol out of everything absorbed and load it onto the lipoproteins that distribute fat around the body. The other seven forms — including gamma — are not selected by α-TTP, so they are largely metabolised and excreted within days. The result:
- α-tocopherol is the form that accumulates and persists in your blood and tissues. This is why nutrition officially defines "vitamin E" as α-tocopherol, why the RDA is set in α-tocopherol, and why supplement labels list α-tocopherol.
- γ-tocopherol and the rest are cleared quickly — even though, as we'll see, gamma may do some of the most valuable anti-inflammatory work.
This sets up the central modern debate, which the bioenergetic commentators (anabology, Hans Amato, and others) have seized on: the body's preference for alpha, and the supplement industry's focus on alpha, may have caused us to throw away the fractions that matter most for chronic disease.
Natural vs synthetic, and bioavailability
A second source of confusion is natural vs synthetic alpha-tocopherol, and it genuinely matters:
- Natural α-tocopherol is labelled "d-alpha-tocopherol" or "RRR-α-tocopherol." It is a single, specific 3-dimensional shape.
- Synthetic α-tocopherol is labelled "dl-alpha-tocopherol" or "all-rac-α-tocopherol." It is a lab mixture of eight different stereoisomers (mirror-image and rotational variants), only some of which the body can use well.
Because α-TTP — the selective liver protein above — strongly prefers the natural RRR shape, natural vitamin E is roughly twice as bioavailable as synthetic for raising tissue levels. Synthetic dl-alpha is cheaper and ubiquitous in cheap multivitamins, and it is the form the bioenergetic critics dismiss as "useless": it is one fraction (alpha), in a partly-unusable synthetic form, with none of the gamma/delta/tocotrienol spectrum.
So when anabology writes "most vitamin E sold today is either synthetic dl-alpha-tocopherol — one fraction, made in a lab — or 'natural' E that's at least half seed oil," this is the precise complaint: the cheap product is the wrong isomer of the wrong single fraction, and even "natural" products are often dilute and carried in PUFA oils.
The mechanism: how vitamin E actually works
This is what the user asked for — not "antioxidant," hand-waved, but how, at the molecular level. Vitamin E's primary job is to stop a specific, dangerous chain reaction called lipid peroxidation, and the way it does it is genuinely elegant.
The problem: lipid peroxidation is a chain reaction
Recall from the gut microbiome and inflammation pages that reactive oxygen species (ROS) attack the polyunsaturated fats (PUFAs) in your cell membranes. Here's why that is so destructive: it is not a one-off. When a radical steals a hydrogen from a PUFA, it turns that fat into a radical, which attacks the next fat, and so on — a self-propagating chain reaction that can rip through an entire membrane, turning structural fat into toxic breakdown products (the 4-HNE and MDA from the gut page). One initial radical can damage hundreds of fat molecules. A membrane undergoing lipid peroxidation is, quite literally, rusting — and the products are inflammatory and mutagenic.
The solution: a chain-breaking antioxidant
Vitamin E is the body's principal lipid-soluble, chain-breaking antioxidant — emphasis on chain-breaking. Sitting embedded in the membrane among the vulnerable fats, when a lipid peroxyl radical forms, vitamin E's chromanol –OH group donates its hydrogen atom to the radical, neutralising it before it can attack the next fat. The chain is broken. One vitamin E molecule stops the whole cascade.
The cost: in giving away its hydrogen, vitamin E itself becomes a radical — the tocopheroxyl radical. But this is a deliberately weak, stable radical: it is not reactive enough to attack the membrane fats, so it sits harmlessly, the chain reaction stopped. Vitamin E, in effect, takes the bullet — sacrificing itself to absorb the damage that would otherwise have propagated. This is why Hans Amato describes it as being "consumed as it neutralizes oxidative stress" — every molecule that does its job is spent.
Regeneration: the antioxidant network
But a spent antioxidant would be wasteful, so the body recycles it — and this is where vitamin E plugs into a whole cooperative network, the mechanism Morph (@doctormorphh) describes. The tocopheroxyl radical can be converted back into active vitamin E by other antioxidants donating an electron to it:
- Vitamin C (ascorbate), working at the membrane–water interface, regenerates vitamin E.
- CoQ10 / ubiquinol (the electron carrier from the electron transport chain) also regenerates it — which is exactly why Morph says taking vitamin E with ubiquinol is "like vitamin E on steroids": ubiquinol "reverses [the tocopheroxyl radical] and makes it active again by donating an electron, turning it back into tocopherol."
flowchart LR
LOO["Lipid peroxyl radical<br/>(chain reaction propagating)"] -->|"Vitamin E donates H<br/>→ CHAIN BROKEN"| STOP[Membrane fat protected]
LOO --> TOXR["Vitamin E becomes<br/>tocopheroxyl radical<br/>(weak, harmless)"]
TOXR -->|"Vitamin C or<br/>CoQ10/ubiquinol<br/>donate an electron"| REGEN["Vitamin E regenerated<br/>(active again)"]
REGEN -->|ready to protect again| LOOSo vitamin E is not a lone soldier — it is the front-line member of a recycling antioxidant team (vitamin E in the membrane, vitamin C at the surface, CoQ10 from the mitochondria), each regenerating the others. This is why isolated antioxidants underperform and why the network matters. It is also the mechanistic core of the "vitamin E kills inflammation" claim: by stopping lipid peroxidation, it cuts off one of the main generators of the inflammatory signals (oxidised-fat damage products) covered on the Inflammation page.
The alpha-vs-gamma debate — why "most vitamin E is useless"
Now the controversy the source posts revolve around. Both anabology and Hans Amato argue that conventional vitamin E (alpha-only) is missing the point, but they emphasise different missing fractions — and the tension between them is illuminating.
The gamma-tocopherol case (anabology, and the Saldeen review). anabology repeatedly cites a 2005 review, "Importance of tocopherols beyond α-tocopherol" (Saldeen & Saldeen), whose argument is:
- γ-tocopherol has anti-inflammatory activity that α-tocopherol lacks. Chemically, gamma's unmethylated 5-position lets it trap reactive nitrogen species (like peroxynitrite) that alpha cannot — and its metabolite inhibits COX-2 (an inflammatory enzyme from the inflammation page).
- Mixed tocopherols (γ + δ + α) outperform alpha alone for antioxidant and anti-inflammatory effect — the review describes a 5:2:1 γ:δ:α preparation as superior.
- High-dose alpha-tocopherol can actually crowd out gamma — supplementing large amounts of alpha lowers blood gamma-tocopherol (they compete for the same handling), so an alpha-only supplement can reduce your protective gamma. The review even notes alpha can be mildly prooxidant in some conditions, with gamma/delta counteracting it.
This is why anabology's product is deliberately gamma-dominant (53 mg γ vs 11 mg α in their formula) and why the slogan is "mixed tocopherols >>" and "vitamin E supplements only list the α-tocopherol content… which is useless [for chronic disease]."
The alpha-tocopherol case (Hans Amato). Hans Amato, coming from a hormonal-optimisation angle, makes almost the opposite emphasis, and the reconciliation is the key insight. He sorts vitamin E products into three categories:
- Alpha-tocopherol isolates — "suppress gamma-tocopherol, create imbalanced signalling, feel disappointing because they're incomplete."
- Wheat germ oil — "too low in concentration, requires large volumes, brings a heavy PUFA load."
- High-gamma products — "protect against inflammation but do nothing for steroidogenesis or testicular function."
His point: gamma is the anti-inflammatory fraction, but alpha is the fraction with the hormonal/steroidogenic effects — and the two jobs are different. For his specific goal (running 1.2 g/week testosterone with no aromatase inhibitor, testing whether vitamin E could blunt aromatisation), he wanted alpha-tocopherol-dominant material — "the closest thing to what [bodybuilder] Vince Gironda used" — mega-dosed at 1500 IU because "vitamin E embeds into membranes and accumulates in endocrine tissue… low doses don't register under high inflammation, high training load, and supraphysiologic androgen levels."
The synthesis: these two aren't contradicting each other — they're describing two different jobs of the vitamin E family. Gamma (and delta) = the anti-inflammatory, lipid-peroxidation-fighting fractions; alpha = the fraction the body retains and that carries the hormonal/membrane-stabilising effects. The practical conclusion both share: avoid synthetic dl-alpha isolates, and favour full-spectrum mixed tocopherols — so you get gamma's anti-inflammatory power and alpha's retained hormonal effects, without crowding either out.
Tocotrienols — the fourth-family bonus
The tocotrienols (the unsaturated-tail half of the family) are less discussed but have their own evidence, which is why questmoosa's hair thread features them:
- Cholesterol-lowering — tocotrienols inhibit HMG-CoA reductase (the same enzyme statins target), modestly lowering cholesterol.
- Hair — the 2010 study questmoosa cites (tocotrienols + alpha, 100 mg/day, 8 months) reported a 34.5% increase in hair count vs a slight decrease on placebo — plausibly via scalp antioxidant protection reducing the oxidative stress around follicles. (As Moosa himself fairly notes, the study had limitations — small, and the effect size should be held loosely — but it's a real signal and points at tocotrienols specifically.)
- Neuroprotection — α-tocotrienol has protective effects on neurons at concentrations alpha-tocopherol doesn't reach.
The takeaway: a truly "full-spectrum" vitamin E includes tocotrienols, not just the four tocopherols — another reason single-fraction products under-deliver.
Vitamin E in the Ray Peat / bioenergetic framework
Now the framework the user asked about. In the bioenergetic world, vitamin E is not a minor antioxidant — it is one of the most important protective molecules, and its role connects directly to the recurring Peat themes (PUFA, estrogen, inflammation, thyroid). The logic:
1. It defends against PUFA — the central Peat irony. The bioenergetic framework's signature claim is that polyunsaturated fats (PUFA) are uniquely damaging because they peroxidise easily (the testosterone deep dive and gut pages). Vitamin E is the body's main defence against exactly that peroxidation. So there's a tight relationship: the more PUFA in your tissues, the more vitamin E you burn through protecting it — your vitamin E requirement rises with PUFA intake. Peat's framing is that vitamin E is the patch for a problem PUFA created; ideally you lower PUFA and keep vitamin E high to neutralise the PUFA already stored in your tissues (which takes years to turn over). This is why vitamin E is a staple "damage-control" supplement in that world.
2. It is anti-estrogen and anti-aromatase. Vitamin E has documented anti-estrogenic activity and can modestly inhibit aromatase (the hormones page enzyme that converts testosterone to estrogen). In the Peat framework, where excess estrogen is treated as inflammatory and anti-metabolic, this makes vitamin E a tool to lower estrogen load — which is exactly why it appears on the Testosterone Kabbalah's list of estrogen-lowering levers, and why Hans Amato tested it as a non-drug aromatase inhibitor under heavy androgen load.
3. It lowers prolactin and supports steroidogenesis. Consistent with its fertility-factor origins, vitamin E (alpha especially) supports the gonadal steroid machinery and can help lower prolactin (the reproductive-suppressing hormone from the hormones page) — fitting the bioenergetic goal of keeping prolactin and estrogen low and androgens functional.
4. It is broadly anti-inflammatory. Via the gamma fraction's COX-2 inhibition and the whole family's lipid-peroxidation braking, vitamin E reduces the oxidative/inflammatory load that the framework sees as the upstream driver of low metabolism, low thyroid, and low testosterone. This is the sense in which the user is exploring it — "a powerful way to kill inflammation while you solve the root cause." That framing is apt: vitamin E is genuinely a symptom-and-damage control (it stops peroxidation and inflammation downstream) while you remove the root cause (lower the PUFA, heal the gut, fix the metabolism).
5. Topical use. anabology's skin products (and Morph's commentary) extend this to the skin: vitamin E protects the skin's lipid barrier from oxidation, which is why it's "the best thing that ever happened to my skin" — though the complaint is that most topical vitamin E products are "half seed oils or terribly sticky," reintroducing the PUFA problem they're meant to solve.
Practical synthesis
Pulling the whole picture together into what actually matters:
- Form matters more than dose. Prefer natural, full-spectrum mixed tocopherols (and ideally tocotrienols) over synthetic dl-alpha isolates. The label should show mixed or d-gamma/d-delta content, not just dl-alpha.
- Take it with fat (it's fat-soluble) and ideally alongside its network partners — vitamin C and CoQ10/ubiquinol regenerate it (Morph's "vitamin E on steroids" point).
- It pairs with lowering PUFA, not replacing it. Vitamin E protects PUFA from peroxidising; it is damage control, most valuable while you reduce dietary PUFA and let stored PUFA turn over.
- Dose is contested. Conventional nutrition uses ~15 mg/day (≈22 IU); the bioenergetic/therapeutic tradition uses far more (100s of IU; Hans Amato's 1500 IU is an extreme, short-term, context-specific experiment, not a baseline recommendation). High-dose alpha-only for long periods is exactly what can crowd out gamma — another argument for mixed tocopherols if dosing high.
⚖️ Calibration. The mechanism (chain-breaking antioxidant, regeneration network) and the alpha-vs-gamma chemistry are solid, mainstream biochemistry. The therapeutic claims — aromatase inhibition, prolactin lowering, the 1500 IU experiment, the hair study — are a mix of plausible mechanism, small studies, and self-experiment, and should be held more loosely. Note too that large trials of high-dose synthetic alpha-tocopherol alone have sometimes shown no benefit or slight harm (e.g. for prostate cancer and all-cause mortality in some meta-analyses) — which the gamma-crowding-out mechanism may partly explain, and which is itself an argument against mega-dosing the isolated synthetic alpha that most of those trials used. "Full-spectrum, natural, modest-to-moderate dose, with the antioxidant network" is the defensible position; "mega-dose dl-alpha forever" is not.
Related compounds & foundations
Vitamin E and its network partners
- Vitamin E — the compound page.
- CoQ10 / ubiquinol — regenerate spent vitamin E (the "vitamin E on steroids" synergy).
- Vitamin C — regenerates vitamin E at the membrane surface.
- Glutathione / NAC — the broader antioxidant network vitamin E plugs into.
- Selenium — cofactor for glutathione peroxidase, which works alongside vitamin E against peroxidation.
Related hormonal / anti-inflammatory tools
- Aspirin — another anti-inflammatory / anti-aromatase tool in the same framework (its own deep dive).
- Progesterone — the anti-estrogen steroid vitamin E complements.
- Fish oil — note the tension: omega-3 is anti-inflammatory but is itself a PUFA that raises vitamin E demand.
Related foundations & deep dives
- Inflammation — lipid peroxidation as an inflammatory driver; what vitamin E interrupts.
- Cellular Energy — membranes, mitochondria, and the CoQ10 that regenerates vitamin E.
- Hormones & the Endocrine System — aromatase, estrogen, and prolactin, which vitamin E modulates.
- Gut Microbiome — the PUFA peroxidation / 4-HNE / MDA damage vitamin E prevents.
- The Testosterone Kabbalah — where vitamin E appears as an estrogen-lowering, anti-inflammatory lever.
Sources informing this page include the peer-reviewed review "Importance of tocopherols beyond α-tocopherol: evidence from animal and human studies" (Saldeen & Saldeen, 2005), and public commentary/threads from @anabology (skin.food), @HansAmato, @doctormorphh, and @questmoosa. Mechanistic and bioavailability claims reflect standard biochemistry; therapeutic and hormonal claims are flagged where they rest on smaller or self-experimental evidence.