Hormones & the Endocrine System
What a hormone actually is, the handful of rules every hormone obeys, and the steroid system in depth — the production tree, free vs bound, DHT and "the alpha enzyme", why testosterone turns into oestrogen, the Ray Peat reframing of oestrogen/progesterone/pregnenolone, and exactly how exogenous hormones shut down your own production.
Why this page exists
Most people carry a cartoon of hormones: testosterone = male, oestrogen = female, and that's roughly it. The reality is a precise, interconnected control system with shared rules, a branching production tree where one hormone becomes another, carrier proteins, receptors, feedback loops, and a surprising sensitivity to your psychological state. This page builds that system from the definition of "hormone" up to the questions that actually puzzle people: why "free" testosterone matters, why extra testosterone can raise oestrogen, what the "5-alpha" everyone mentions is, why the Ray Peat world treats oestrogen as a villain and progesterone as a hero, and how taking a hormone shuts off your own supply.
It is the chemical counterpart to the autonomic nervous system page: where the ANS is the body's fast, electrical, point-to-point control, the endocrine system is the slow, chemical, broadcast control. The two together run everything. We've already met pieces of it — the thyroid and cortisol axes on the metabolism page, the liver's hormone clearance and the gut's estrobolome. This page covers the steroid (sex) hormones in depth and, more importantly, extracts the shared rules that all hormones obey — so the whole system becomes one pattern rather than a list of facts.
What a hormone actually is
A hormone is a chemical messenger: a molecule released by one tissue that travels (usually through the blood) to act on distant target cells, changing their behaviour. That word distant is the key to the definition — it distinguishes a hormone from a neurotransmitter (which acts across a tiny synaptic gap) and from local signals (which act on neighbours). A hormone is broadcast: released into the bloodstream and carried body-wide, to act wherever there are cells equipped to receive it.
That last clause is the second half of the definition, and it is the single most important concept on this page: a hormone does nothing on its own. It only has an effect where there is a receptor for it.
A receptor is a protein, on or inside a target cell, shaped to recognise and bind one specific hormone — a lock to the hormone's key. The hormone circulates everywhere, but it only acts on cells that display its receptor. This is why the same testosterone molecule builds muscle in muscle tissue, grows a beard on the face, and does nothing at all to a kidney cell — the difference is which cells carry the androgen receptor and how many. The hormone is the message; the receptor is the ear. No receptor, no effect, no matter how much hormone.
This gives us the master equation for any hormone's real-world effect:
Effect = hormone level × number of receptors × sensitivity of those receptors
Hold onto this. It answers one of the deepest questions about hormones — whether a problem is too little hormone, too few receptors, or desensitised receptors — and we return to it at the end.
The one big split: two kinds of hormone, two mechanisms
Here is the unifying framework. Despite their variety, essentially all hormones fall into one of two structural classes, and the class dictates everything about how the hormone behaves — how it travels, how fast it acts, how long it lasts, and what kind of receptor it uses.
Class 1: Water-soluble hormones (peptides and amines — e.g. insulin, adrenaline, most pituitary hormones like LH, FSH, prolactin, growth hormone).
- They dissolve freely in blood (which is watery), so they travel easily on their own.
- They cannot cross the cell membrane (which is fatty, and repels water-soluble molecules — recall this barrier from the cellular energy and digestion pages). So they must bind a receptor on the cell's outer surface.
- That surface receptor triggers a relay of internal "second messengers" — a fast cascade inside the cell. The effect is fast (seconds to minutes), short-lived, and amplifying. Adrenaline making your heart pound within a second is this class in action.
Class 2: Fat-soluble hormones (steroids — testosterone, oestrogen, progesterone, cortisol — and the thyroid hormones).
- They do not dissolve well in blood, so they must be carried on transport proteins (the basis of "free vs bound" — next section).
- But because they are fat-soluble, they pass straight through the cell membrane into the cell.
- Inside, they bind intracellular / nuclear receptors — receptors in the cytoplasm or nucleus — and the hormone-receptor pair then acts directly on the DNA, switching genes on or off (it becomes a transcription factor, exactly as T3 did on the metabolism page). Because this works by changing which proteins the cell builds, the effect is slow (hours to days), long-lasting, and profound.
flowchart TD
subgraph WS["WATER-SOLUBLE (peptides, amines)<br/>insulin · adrenaline · LH · prolactin"]
W1[Travels free in blood] --> W2[Binds SURFACE receptor<br/>can't enter cell] --> W3["Second-messenger cascade<br/>→ FAST, short-lived"]
end
subgraph FS["FAT-SOLUBLE (steroids + thyroid)<br/>testosterone · oestrogen · cortisol"]
F1[Rides carrier protein in blood] --> F2[Passes THROUGH membrane<br/>binds nuclear receptor] --> F3["Changes gene transcription<br/>→ SLOW, long-lasting"]
end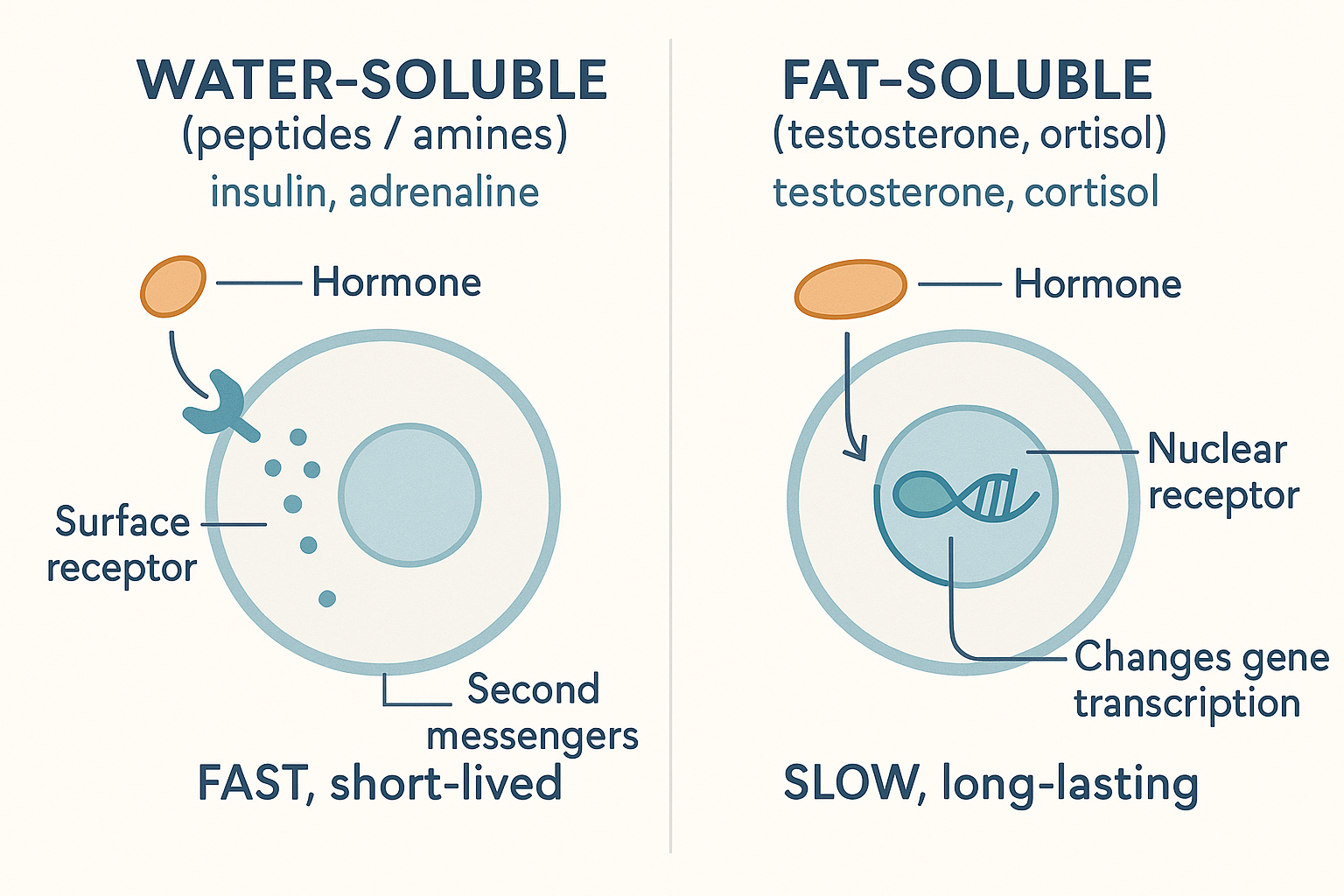 The master split: water-soluble hormones knock on the door (surface receptor, fast); fat-soluble steroids walk in and rewrite the instructions (nuclear receptor, slow and lasting).
The master split: water-soluble hormones knock on the door (surface receptor, fast); fat-soluble steroids walk in and rewrite the instructions (nuclear receptor, slow and lasting).
This single split explains an enormous amount, and it directly answers the question "do all hormones follow common patterns?" — yes: they all follow the pattern produce → travel → bind a receptor → cause an effect → get cleared, but they divide into these two mechanistic families. The rest of this page is mostly about Class 2, the steroids — but the framework is universal.
How hormones travel: free vs bound (and why it applies to all the carried ones)
Because the fat-soluble hormones can't dissolve in blood, they ride on carrier proteins. The main ones:
- SHBG (sex-hormone-binding globulin) — carries testosterone and oestrogen.
- Albumin — a general-purpose carrier (the liver-made protein from the liver page) that loosely carries many hormones.
- CBG (cortisol-binding globulin) — carries cortisol.
- Thyroid-binding globulin — carries thyroid hormone.
Here is the crucial rule: only the free (unbound) fraction of a hormone is biologically active. A hormone molecule clamped onto a carrier protein cannot enter cells or bind receptors — it is in transport/storage, inert until released. Only the small percentage floating free can actually act.
This is the answer to "free vs bound testosterone," and to the user's question of whether it applies to all hormones: yes — free vs bound applies to every carried (fat-soluble) hormone, not just testosterone. And it has large practical consequences:
- A man can have "normal total testosterone" on a blood test but be functionally low because most of it is bound to SHBG and little is free. Total tells you the inventory; free tells you what's actually usable.
- Anything that raises SHBG (high oestrogen, certain thyroid states, ageing, some medications) lowers free hormone even if total is unchanged — a hidden way to become functionally deficient.
- This is a third lever on the master equation: even before receptors, the binding proteins decide how much hormone is available to act at all.
flowchart LR
TOTAL[Total testosterone in blood] --> BOUND["BOUND to SHBG/albumin<br/>(inactive — storage/transport)"]
TOTAL --> FREE["FREE (unbound)<br/>~1–2% — the ONLY active fraction"]
FREE --> ACT[Enters cells, binds receptors,<br/>has an effect]
BOUND -.->|cannot act| X[No effect until released]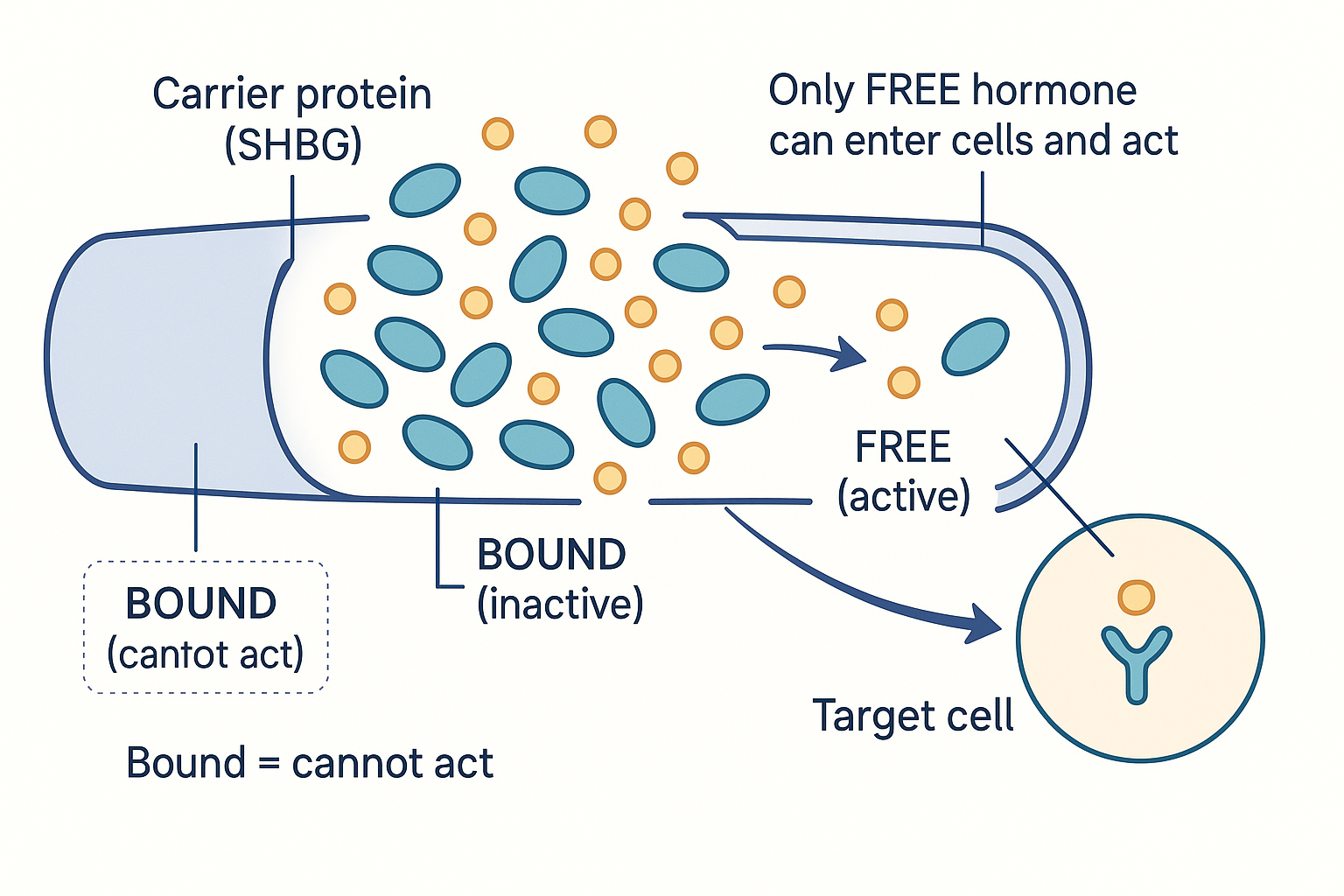 Only the free fraction acts: most hormone rides bound to carriers like SHBG, inert until released — which is why "normal total" can still mean "low usable."
Only the free fraction acts: most hormone rides bound to carriers like SHBG, inert until released — which is why "normal total" can still mean "low usable."
Where hormones are made, and where they go
Made — by the endocrine glands, the dedicated hormone factories: the hypothalamus and pituitary (the brain's control glands), the thyroid (metabolic tempo), the adrenals (cortisol, adrenaline, DHEA), the gonads (testes → testosterone; ovaries → oestrogen and progesterone), the pancreas (insulin), and the pineal (melatonin). Crucially, some hormones are also made by tissues you wouldn't call glands: fat tissue is a major endocrine organ — it makes oestrogen (via aromatase, below) and leptin — which is why body fat directly shapes the hormonal picture.
Cleared — a hormone's signal has to end, or control is impossible. Three mechanisms remove it: (1) receptor binding and internalisation consumes some; (2) the liver inactivates hormones via the Phase I/II detox machinery from the liver page (tagging them for excretion) and dumps them into bile or urine; (3) the gut can recycle some (the estrobolome reactivating oestrogen, from the gut microbiome page). How long a hormone lasts before clearance is its half-life — seconds for adrenaline, hours to days for steroids — which tracks the water-soluble/fat-soluble split again.
Why hormones at all — why have this slow chemical system when nerves are faster? Because some jobs need slow, sustained, body-wide broadcast: growth over years, the reproductive cycle, the day-long arc of cortisol, the lifelong programme of puberty. Nerves are fast but point-to-point; hormones are slow but reach every cell at once and hold a state for hours or years. The two systems are complementary — and the hypothalamus sits at the junction, translating between them.
The steroidogenesis tree: how one hormone becomes another
Now the diagram people see and find bewildering — the branching map where hormones turn into other hormones. It is called steroidogenesis (making steroids), and once you see the logic it is straightforward: every steroid hormone is built from cholesterol, step by step, and each arrow is a single enzyme. Hormones "turn into" each other because they are stages along a production line.
The trunk of the tree:
- Cholesterol is the raw material for all steroid hormones. (This is one reason cholesterol is not simply "bad" — it is the substrate for every sex and stress hormone you have.)
- Cholesterol → Pregnenolone. This is the rate-limiting first step, happening inside mitochondria, and it is the gateway to the entire tree. Pregnenolone is therefore called the "mother hormone" — everything downstream flows from it.
- From pregnenolone the tree branches:
- one branch → progesterone → (onward) → cortisol (the stress branch) and aldosterone (salt/blood-pressure branch);
- another branch → DHEA → androstenedione → testosterone (the androgen branch).
- And testosterone itself sits at a final fork:
- Testosterone → DHT via the enzyme 5-alpha-reductase (the potent-androgen branch);
- Testosterone → oestrogen via the enzyme aromatase (the oestrogen branch).
flowchart TD
CHOL[Cholesterol] -->|"rate-limiting step"| PREG["Pregnenolone<br/>(the 'mother hormone')"]
PREG --> PROG[Progesterone]
PREG --> DHEA[DHEA]
PROG --> CORT["Cortisol<br/>(stress)"]
PROG --> ALDO["Aldosterone<br/>(salt/BP)"]
DHEA --> ANDRO[Androstenedione]
ANDRO --> TEST[Testosterone]
TEST -->|"5-alpha-reductase"| DHT["DHT<br/>(most potent androgen)"]
TEST -->|"aromatase"| EST["Oestrogen<br/>(oestradiol)"]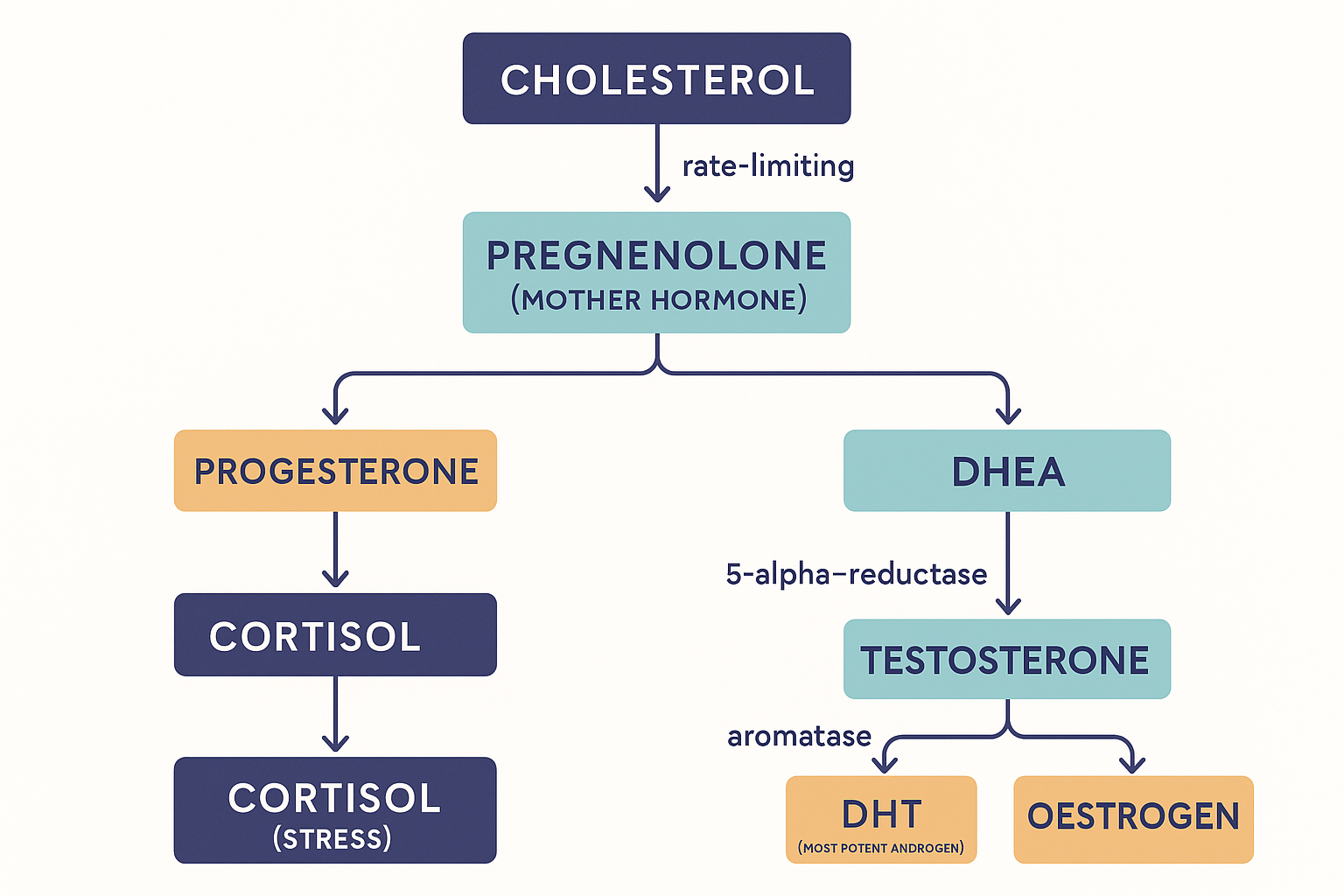 Every steroid is built from cholesterol down this tree, each arrow an enzyme — which is why testosterone is one enzyme from the most potent androgen (DHT) and a different enzyme from oestrogen.
Every steroid is built from cholesterol down this tree, each arrow an enzyme — which is why testosterone is one enzyme from the most potent androgen (DHT) and a different enzyme from oestrogen.
Two features of this tree explain most of the puzzles:
- The branches share precursors and compete. Because pregnenolone and progesterone feed both the cortisol branch and the sex-hormone branches, demand on one branch can affect the others — the basis of the "pregnenolone steal" idea below.
- The same starting hormone can become opposite things depending on which enzyme acts. Testosterone is one enzyme away from the most potent androgen (DHT) and one different enzyme away from oestrogen. Which way it goes depends on which enzymes are active in which tissue — which is why the same hormone has such different fates around the body.
The whole tree depends on cofactors and conditions: cholesterol as substrate, adequate thyroid hormone and vitamin A to run steroidogenesis efficiently (a Peat-favoured point), plus the usual enzyme cofactors. A sluggish thyroid or low cholesterol can throttle the entire tree at the top.
DHT and 5-alpha-reductase: "the alpha" you keep hearing about
The "alpha" in "5-alpha-reductase" and "alpha-reductase inhibitors" refers to this enzyme: 5-alpha-reductase, which converts testosterone → DHT (dihydrotestosterone).
DHT is the answer to "the most potent male hormone despite not being the most abundant." It circulates at much lower levels than testosterone, but it binds the androgen receptor far more strongly and more durably — roughly several times the potency. So in the tissues where 5-alpha-reductase is active, DHT, not testosterone, is the dominant androgen signal. Those tissues are the skin and hair follicles, the prostate, and the external genitals — which is why DHT drives beard growth, body hair, prostate growth, and (in genetically susceptible follicles) male-pattern baldness.
This is why 5-alpha-reductase inhibitors exist. Drugs like finasteride block the enzyme, lowering DHT — used for hair loss and enlarged prostate. But because DHT has roles beyond hair and prostate (libido, mood, erectile function), blocking it can carry sexual and mood side effects in some people. It is a clean illustration of the tree's logic: inhibit one enzyme (one arrow) and you redirect the whole downstream flow, with both intended and unintended consequences.
Aromatase: why "extra" testosterone can become oestrogen
The other fork off testosterone is aromatase, the enzyme that converts androgens (testosterone) into oestrogen. This answers the user's puzzle directly: yes — testosterone is routinely converted to oestrogen, and more so when there is more substrate or more aromatase activity.
The critical fact: aromatase is highly active in fat tissue. So:
- More body fat → more aromatase → more conversion of testosterone to oestrogen → lower testosterone and higher oestrogen simultaneously. This is a self-reinforcing trap: higher body fat lowers the androgen/oestrogen ratio, which tends to promote more fat storage. It's a major reason body composition and hormones are so entangled in men.
- This is why simply pushing testosterone up (e.g. with exogenous testosterone) can raise oestrogen too — the excess testosterone is substrate for aromatase. Bodybuilders therefore often pair testosterone with aromatase inhibitors to limit this conversion (with their own risks, since some oestrogen is necessary).
So "extra testosterone converting to oestrogen" is not a malfunction — it is the aromatase arm of the tree doing exactly what it does, scaled up by substrate availability and by how much fat tissue (aromatase) is present.
Oestrogen, progesterone, pregnenolone — the Ray Peat reframing
The biohacking discourse you've encountered is heavily shaped by Ray Peat's framework, which reframes the sex steroids in a way that is worth presenting clearly (and with the note that it is a perspective — internally coherent and mechanistically argued, but more heterodox than mainstream endocrinology, which weights things differently).
Oestrogen — not "the female hormone," and excess is the concern. In the Peat view, oestrogen is not a "female essence" but one signalling molecule with a specific character: it is proliferative (drives cell division — relevant to some cancers), pro-inflammatory in excess, water-retaining, and associated with the stress and ageing state. Everyone of both sexes has and needs some; the concern is excess ("oestrogen dominance"), which is promoted by body fat (aromatase, above), by impaired liver clearance and the gut estrobolome recycling it (gut microbiome page), and by stress. This is why the Peat-influenced community treats lowering excess oestrogen — rather than worshipping testosterone — as a central goal.
Progesterone — the protective counter-hormone. Despite its reputation as a "female/pregnancy hormone," progesterone in this framework is a broadly protective, anti-oestrogen, pro-thyroid, calming hormone for both sexes. Two mechanisms stand out: it directly opposes oestrogen's proliferative effects, and its metabolite allopregnanolone is a potent positive modulator of the GABA-A receptor — the brain's main calming system (from the ANS and gut-brain pages) — which is why progesterone is calming and sleep-promoting. Hence its high regard in that community.
Pregnenolone — the "mother" and anti-stress reserve. Because pregnenolone sits at the top of the tree (the mother hormone), the Peat argument is that keeping it abundant gives the body raw material to make whatever downstream hormone it needs — and that it has its own protective, pro-cognitive, anti-stress character. This connects to the famous "pregnenolone steal" (or "cortisol steal") idea: under chronic stress, the body's heavy demand for cortisol is thought to divert the shared upstream precursors (pregnenolone, progesterone) toward the cortisol branch, at the expense of the protective/sex-hormone branches (DHEA, progesterone, testosterone). The picture: chronic stress = high cortisol, low DHEA/progesterone — "stress steals from your protective hormones." (A caveat for honesty: strict biochemists note that steroid synthesis is compartmentalised within each gland, so a literal shared-pool "steal" is debated; but as a systems description of the stress-driven shift in hormone output, the pattern is real and observable.) This is the rationale for supplementing pregnenolone or progesterone in that framework — topping up the protective branch the stress system depletes.
Prolactin: useful in its place, harmful in excess
Prolactin is a water-soluble pituitary hormone, best known for driving milk production — but it does much more, and the user's instinct ("not good, but necessary, and good before bed") is exactly right. The nuances:
- Chronically high prolactin is harmful. It suppresses the sex-hormone axis (lowering testosterone and oestrogen), kills libido, and is associated with the stress state. Persistently elevated prolactin is a recognised cause of low libido and reproductive problems in both sexes.
- It is regulated by dopamine. Here is the link to the cognition page: dopamine suppresses prolactin. High dopamine (motivated, driven, post-reward states) keeps prolactin low; low dopamine lets prolactin rise. This is part of why the post-orgasm drop in drive (the refractory period) involves a prolactin surge, and why dopamine-supporting states feel anti-prolactin.
- It naturally rises at night, and that's appropriate. Prolactin has a strong circadian rhythm, peaking during sleep. This nocturnal rise is part of the normal rest-and-repair state — so "good before bed" is correct: a night-time prolactin rise accompanies healthy sleep and recovery. The problem is daytime/chronic elevation, not the natural nocturnal peak. Context is everything: the same hormone is part of healthy sleep at night and a libido-killing stress signal when chronically high by day.
The feedback system: how the body senses and controls hormone levels
Now the engineering question the user asked precisely: what sensor detects hormone levels, what mechanism feeds that back, and how does exogenous hormone cause shutdown? The answer is a negative feedback axis, identical in architecture to the thyroid (HPT) and stress (HPA) axes from the metabolism page. For the sex hormones it is the HPG axis (hypothalamus–pituitary–gonad):
- The hypothalamus releases GnRH (gonadotropin-releasing hormone) in pulses.
- The pituitary responds by releasing LH and FSH into the blood.
- The gonads respond: in men, LH tells the testes' Leydig cells to make testosterone (FSH supports sperm production); in women, LH/FSH drive the ovarian cycle and oestrogen/progesterone.
- The feedback (the "sensor"): the circulating testosterone and oestrogen travel back up to the hypothalamus and pituitary, which detect them (via their own receptors) and, when levels are high enough, reduce GnRH and LH/FSH output. The output inhibits its own command signal — a thermostat. This is the sensor and the retro-feed: the hypothalamus/pituitary are continuously measuring the circulating hormone and throttling production to hold it in band.
flowchart TD
HYP[Hypothalamus] -->|GnRH| PIT[Pituitary]
PIT -->|LH / FSH| GON[Gonads]
GON -->|Testosterone / Oestrogen| BODY[Body effects]
GON -.->|"negative feedback:<br/>'levels are high, ease off'"| PIT
GON -.->|negative feedback| HYP
EXO[Exogenous hormone<br/>injected/supplemented] -.->|"feedback reads 'plenty'"| PIT
EXO -.-> HYP
PIT -->|"LH/FSH shut off"| SHUT["Gonads stop producing<br/>→ testicular shrinkage"]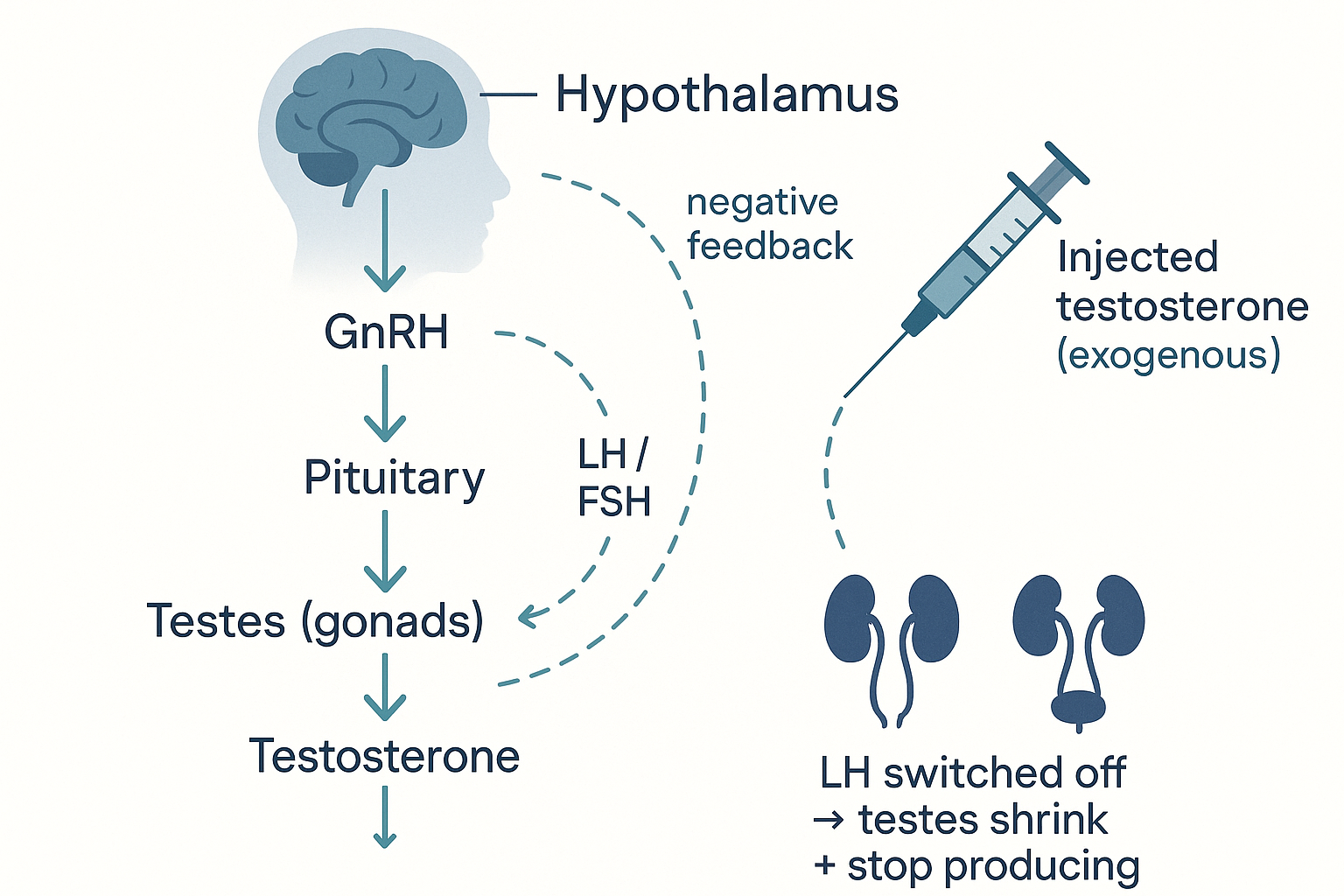 The feedback axis is the sensor: the hypothalamus and pituitary measure circulating hormone and throttle their signals — so injected hormone reads as "plenty," shuts off LH, and idles your own factory.
The feedback axis is the sensor: the hypothalamus and pituitary measure circulating hormone and throttle their signals — so injected hormone reads as "plenty," shuts off LH, and idles your own factory.
Now the exogenous-hormone shutdown, precisely. When you take an external hormone (e.g. testosterone), the hypothalamus and pituitary sense the high circulating level through that same feedback mechanism — they cannot tell the difference between hormone you made and hormone you took. Reading "we have plenty," they shut down GnRH and LH/FSH. With no LH signal, the testes' Leydig cells stop producing testosterone and shrink (testicular atrophy), and sperm production falls. The body has, in effect, been fooled into switching off its own factory because the warehouse looks full. This is exactly why anabolic steroid use suppresses natural production and fertility.
Recovery: when the exogenous hormone is stopped, circulating levels fall, the feedback brake releases, the hypothalamus resumes GnRH pulses, LH/FSH return, and the gonads are stimulated to start producing again. But recovery is slow (weeks to many months) because the axis has to "wake up" and the gonads have to regain capacity — and it is sometimes incomplete, especially after long or high-dose use, which is why post-cycle recovery protocols exist and why suppression is not always fully reversible. The system is robust but not infinitely so.
Mind into hormone: why perception changes your endocrine output
The user's most striking question: how can something experienced psychologically — the environment, social status, a feral pig's wild surroundings — physically change hormone production so strongly? The answer follows directly from where the axis starts: at the top sits the hypothalamus, which is part of the brain.
The hypothalamus does not operate in a vacuum — it receives heavy input from the limbic system, including the amygdala (threat/emotion) and the brain's appraisal of social status, competition, safety, and stress. Because the hypothalamus is the first link in the HPG (and HPA) axis, whatever the brain concludes about the environment is translated directly into hormone output. Concretely:
- Winning, dominance, and competition raise testosterone; losing and subordination lower it — measurably, within minutes to hours (the "winner effect," documented in humans and animals). The brain reads social outcome and adjusts androgen output.
- Chronic stress raises cortisol, which suppresses GnRH (and the whole HPG axis), lowering testosterone — the endocrine signature of feeling chronically threatened.
- Environmental and social context — including, in animals, the shift from domesticated to feral living — changes androgen output via this brain-to-gonad route. A domestic animal returned to a wild, competitive, free environment experiences a different stream of social and threat signals, the hypothalamus reads "different world," and androgen output (and androgen-driven traits like the DHT-dependent features) shifts accordingly. The "psyche changing the body" is not mystical: the brain's appraisal of the environment is literally the input to the hormone-control axis.
This is profound and worth sitting with: your endocrine system is downstream of your perception. The same machinery means that lifestyle, status, stress, sleep, and even mindset measurably move hormone levels — because they all feed the hypothalamus that commands the axis.
Production vs expression vs sensitivity: the unifying payoff
Finally, the user's deepest question: is a hormone issue about production, expression, or sensitivity — and do all hormones share common patterns? Return to the master equation:
Effect = hormone level × number of receptors × sensitivity of receptors (× free fraction)
A "hormone problem" can live at any of these points, and they are genuinely different:
- Production — the gland makes too little (or too much) hormone. (e.g. failing testes, or an over-active tumour.)
- Transport / free fraction — enough hormone is made, but too much is bound to carrier proteins, so too little is free and active. (High SHBG masking "normal" total testosterone.)
- Receptor expression — enough free hormone, but the target cells display too few receptors to hear it. (Often regulated down by chronic over-exposure.)
- Receptor sensitivity — receptors are present but desensitised — they no longer respond well. (Exactly what insulin resistance is, from the metabolism page: plenty of insulin, plenty of receptors, but a jammed response.)
This is why two people with identical blood hormone levels can have completely different symptoms — the levels are only one of four terms. It also reframes treatment: sometimes the fix is more hormone, but often it is fewer carrier proteins, more receptors, or better sensitivity — and chronically flooding a system with hormone tends to down-regulate its own receptors, making the sensitivity problem worse (the same trap as caffeine tolerance and insulin resistance).
And it answers the unifying question: yes, all hormones share the same underlying pattern — produced by a gland → carried in blood (free vs bound) → bind a receptor (surface or nuclear) → cause an effect → get cleared → with the whole thing held in check by negative feedback. Whether it's insulin, cortisol, thyroid, testosterone, or oestrogen, that skeleton is identical. The differences — fast vs slow, surface vs nuclear receptor, which gland, which feedback axis — are variations on this one template. Learn the template and every hormone becomes a special case of it rather than a separate fact to memorise.
Regulation at a glance: what drives each hormone up and down
Everything above explains the machinery. But when people actually reason about hormones, they think in terms of "what raises this, what lowers it, and what does it then do?" — the kind of driver-graph the Testosterone Kabbalah deep dive is built around. Here are those graphs for the major hormones, as quick-reference maps. Read each one as: drivers (left) → the hormone (centre) → what it does (right). They are deliberately simplified — every arrow is a relationship explained somewhere on this page or the linked foundations.
Testosterone
flowchart LR
UP["↑ Raised by:<br/>LH signal · sleep · low body fat ·<br/>zinc · vitamin D · winning/dominance ·<br/>low stress"] --> T((Testosterone))
DOWN["↓ Lowered by:<br/>cortisol/stress · high oestrogen (feedback) ·<br/>high prolactin · endotoxin/inflammation ·<br/>low thyroid · obesity (aromatase) ·<br/>exogenous testosterone (shutdown)"] --> T
T --> EFF["Drives:<br/>muscle · libido · drive ·<br/>→ DHT · bone density"]Oestrogen (oestradiol)
flowchart LR
UP["↑ Raised by:<br/>aromatase in body fat · ample testosterone substrate ·<br/>poor liver clearance · gut β-glucuronidase recycling ·<br/>endotoxin · PUFA · insulin resistance"] --> E((Oestrogen))
DOWN["↓ Lowered by:<br/>fat loss (less aromatase) · good liver + gut ·<br/>vitamin E · progesterone · aromatase inhibitors"] --> E
E --> EFF["Excess drives:<br/>proliferation · inflammation ·<br/>↑ prolactin · ↑ SHBG · water retention<br/>(but some is needed: bone, libido, brain)"]Cortisol
flowchart LR
UP["↑ Raised by:<br/>psychological stress · low blood sugar/fasting ·<br/>poor sleep · inflammation/endotoxin ·<br/>overtraining · low thyroid"] --> C((Cortisol))
DOWN["↓ Lowered by:<br/>adequate carbs + regular meals · salt ·<br/>sleep · sunlight · magnesium · theanine"] --> C
C --> EFF["Drives:<br/>↑ blood glucose (gluconeogenesis) ·<br/>↓ T4→T3 · ↓ GnRH → ↓ testosterone ·<br/>↑ aromatase · visceral fat · immune suppression"]Thyroid (active T3)
flowchart LR
UP["↑ Raised by:<br/>T4→T3 conversion (liver + selenium) ·<br/>adequate carbs/glycogen · iodine · low stress"] --> T3((Active T3))
DOWN["↓ Lowered by:<br/>cortisol/stress · fasting/low-carb ·<br/>inflammation · low selenium ·<br/>high oestrogen · poor liver → reverse T3"] --> T3
T3 --> EFF["Drives:<br/>metabolic rate · body temperature ·<br/>mitochondrial biogenesis · Na/K pumps ·<br/>5α-reductase → DHT"]Insulin
flowchart LR
UP["↑ Raised by:<br/>dietary carbohydrate/glucose · frequent eating ·<br/>insulin resistance (compensatory)"] --> I((Insulin))
DOWN["↓ Lowered by:<br/>fasting · low-carb · exercise · fat loss"] --> I
I --> EFF["Drives:<br/>glucose INTO cells · fat STORAGE ·<br/>blocks lipolysis (no fat-burning) ·<br/>glycogen + protein synthesis"]Prolactin
flowchart LR
UP["↑ Raised by:<br/>serotonin · oestrogen · stress · poor sleep ·<br/>dopamine suppression · frequent ejaculation ·<br/>low zinc"] --> P((Prolactin))
DOWN["↓ Lowered by:<br/>dopamine · sleep · zinc · vitamin E ·<br/>P5P (B6) · low oestrogen/serotonin"] --> P
P --> EFF["Excess drives:<br/>↓ GnRH → ↓ testosterone/DHT · ↓ libido<br/>(the natural night-time rise aids sleep/recovery)"]Bonus: chained regulation — the 'never goon' example
These driver-graphs also chain together, which is how the biohacking community reasons about knock-on effects. A widely-shared example (from @BerbarianWizard and replies) traces frequent ejaculation through several of the graphs above:
frequent ejaculation → ↓ zinc → ↑ copper → ↑ oestrogen → ↑ prolactin → negative feedback at the hypothalamus → ↓ testosterone.
Each link is one of the relationships on this page (zinc is an aromatase cofactor that keeps the copper:zinc ratio in check; oestrogen and prolactin both feed back to suppress GnRH). Whether the magnitude of the effect from normal ejaculation frequency is meaningful is debated — community notes point out research shows no lasting testosterone drop from frequent ejaculation — but it's a clean illustration of how the individual driver-graphs connect into a cascade.
Putting it all together
- A hormone is a chemical messenger that travels (usually in blood) to act on distant cells that carry its receptor — no receptor, no effect. Effect = hormone level × receptor number × receptor sensitivity × free fraction.
- Two structural classes, two mechanisms: water-soluble (peptides/amines) bind surface receptors for fast, brief effects; fat-soluble (steroids + thyroid) enter the cell and bind nuclear receptors, changing gene transcription for slow, lasting effects.
- Fat-soluble hormones ride carrier proteins, and only the free (unbound) fraction is active — this "free vs bound" applies to all the carried hormones (testosterone, oestrogen, cortisol, thyroid), and explains "normal total but low free."
- Steroidogenesis is a tree: all steroids are built from cholesterol → pregnenolone (the rate-limiting "mother hormone") → branching to progesterone/cortisol and to DHEA/testosterone — each arrow an enzyme, which is why hormones "turn into" each other.
- 5-alpha-reductase ("the alpha") converts testosterone → DHT, the most potent androgen despite low levels (skin, hair, prostate); aromatase converts testosterone → oestrogen, especially in fat tissue — which is why more body fat lowers testosterone and raises oestrogen, and why "extra" testosterone can become oestrogen.
- The Ray Peat reframing: excess oestrogen is the concern (proliferative, stress/ageing-linked), progesterone is protective and calming (allopregnanolone → GABA), and pregnenolone is the upstream reserve depleted by chronic stress (the "pregnenolone steal" — high cortisol at the expense of protective hormones).
- Prolactin is necessary and rises healthily at night (rest/repair) but is harmful when chronically high by day (suppresses sex hormones and libido); dopamine suppresses it.
- Negative feedback axes (HPG, like HPT/HPA) are the sensor-and-control: the hypothalamus/pituitary detect circulating hormone and throttle their signals — which is exactly why exogenous hormone shuts down your own production (the feedback reads "plenty," LH/FSH switch off, the gonads idle and shrink), and why recovery on stopping is slow and sometimes incomplete.
- Perception drives hormones: because the hypothalamus (a brain region reading status, stress, and environment) sits atop the axis, social and psychological state measurably shifts hormone output — the real mechanism behind "environment changes the body."
- A hormone problem can be production, free fraction, receptor number, or sensitivity — and chronically flooding a system down-regulates its own receptors. All hormones share the same produce → carry → receptor → effect → clear → feedback template.
The unifying idea: the endocrine system is one template repeated — a messenger, a carrier, a receptor, an effect, and a feedback loop — overlaid on a branching production tree built from cholesterol and governed, at the top, by the brain's reading of your world. Master the template and the steroid tree, and the whole bewildering hormone landscape collapses into a system you can reason about.
Related Compounds & Deep Dives
Sex steroids & the tree
- Testosterone — the principal androgen; substrate for both DHT (via 5-alpha-reductase) and oestrogen (via aromatase).
- DHT — the most potent androgen; dominant in skin, hair, and prostate.
- Oestrogen — reframed as a signalling molecule whose excess is the concern; the estrobolome and aromatase set its load.
- Progesterone — anti-oestrogen, pro-thyroid, calming via allopregnanolone/GABA.
- Pregnenolone — the upstream "mother hormone"; depleted by chronic stress (pregnenolone steal).
Stress & feedback
- Cortisol — the stress branch that competes for precursors and suppresses the HPG axis.
- LH — the pituitary signal that tells the gonads to produce; suppressed by exogenous hormone.
Cofactors & modulators
- Boron — may raise free testosterone (lowers SHBG binding).
- Zinc, magnesium, vitamin D — cofactors supporting healthy steroidogenesis and free-hormone levels.
- Tadalafil — vascular/erectile support (acts downstream of androgens).
Other endocrine signals
- Insulin — the archetypal water-soluble hormone and the clearest example of receptor sensitivity failure (insulin resistance).
- Melatonin — the pineal hormone of circadian timing.
- Growth hormone / IGF-1 — the growth axis.
- Oxytocin — a peptide hormone of bonding and calm.
Related foundations
- Systemic Metabolism — the HPT and HPA axes that share this exact feedback architecture, and the cortisol/insulin systems.
- Liver — where hormones are cleared (Phase I/II) and where T4→T3 conversion happens.
- Gut Microbiome — the estrobolome that recycles oestrogen and shapes its load.
- Autonomic Nervous System — the fast electrical control system the endocrine system complements.