Fat & Fat Metabolism
The most argued-about molecule in nutrition — and the one most people can't actually define. This page builds fat from the atom up: what a fatty acid is, the two-axis map that makes "saturated / monounsaturated / PUFA / trans / MCT" second nature, why fat carries so much energy, exactly how it's burned, why polyunsaturated fat is uniquely prone to setting your cells on fire — and the full bioenergetic cascade behind the claim that carbohydrate, not fat, is the cleaner metabolic fuel.
Read alongside
This page goes deeper on fuel than Cellular Energy (start there for the ATP basics), connects its peroxidation section to the Vitamin E deep dive and the Inflammation page, and sets up the Systemic Metabolism page that follows it. It is intentionally the most detailed foundations page.
Why this page exists
Fat is where nutrition arguments go to die. Is saturated fat bad? Are seed oils poison? Is keto optimal or a stress state? You cannot evaluate any of these claims without first understanding what these molecules actually are — and almost nobody does, because "fat" gets discussed as one undifferentiated blob. It is not. The word covers molecules as different from each other as a diamond is from coal, and once you can see the molecular differences, the metabolic and health arguments stop being a clash of opinions and become legible.
So this page is built in two halves. First, the molecule — we make the fat categories (saturated, monounsaturated, polyunsaturated, MCT, trans) genuinely second nature, by chemistry, shape, digestion, metabolism, and hormonal effect. Then, the system — how fat is burned, why polyunsaturated fat is uniquely dangerous to burn, and the full Ray Peat–influenced cascade behind "carbs are the cleaner fuel." Where the popular framing (including a fact-checked version of a widely-circulated AI explainer) overstates its case, it's flagged ⚖️ Calibration.
Part 1 — What fat actually is
The fatty acid: a head and a tail
Almost all dietary and body fat is built from a single repeating unit: the fatty acid. And a fatty acid has exactly two parts, with opposite personalities:
- The tail — a long chain of carbon (C) atoms bonded to hydrogen (H) atoms. This is the "fatty" part: oily, greasy, water-fearing (hydrophobic). It blends with other fats and refuses to mix with water.
- The head — a small cluster at one end called a carboxyl group, written –COOH (a carbon double-bonded to one oxygen and single-bonded to an –OH). This is the "acid" part: water-loving (hydrophilic).
[ ===== HYDROCARBON TAIL ===== ] [ CARBOXYL HEAD ]
(oily, water-fearing) (acidic, water-loving)
H H H H H H O
| | | | | | //
H — C — C — C — C — C — C — — — — — — — — — — C
| | | | | | \
H H H H H H O—H
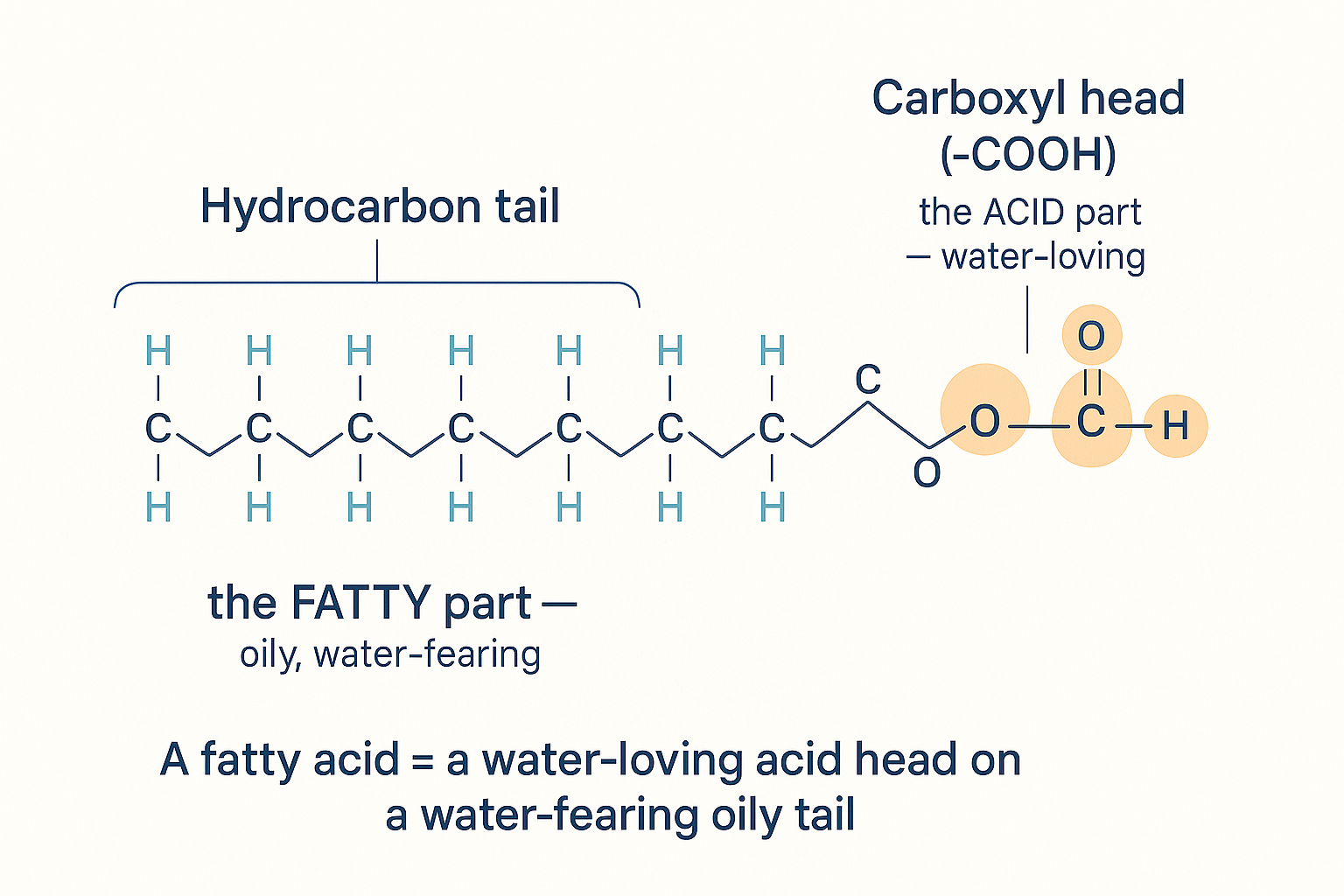 Every fatty acid is this: a water-loving acid head on a water-fearing oily tail. The tail's length and bonds are what vary.
Every fatty acid is this: a water-loving acid head on a water-fearing oily tail. The tail's length and bonds are what vary.
Why "acid"? In chemistry, an acid is anything that readily donates a hydrogen ion (H⁺). That –OH on the carboxyl head lets go of its hydrogen easily, so a fatty acid technically is an acid. But it doesn't taste sour or burn like vinegar, for two reasons: the huge oily tail drowns out the tiny acid head, and because the molecule won't dissolve in water (saliva), the acid head stays "hidden." The shorter the tail, the less camouflage — which is why acetic acid (a 2-carbon fatty acid) is sour vinegar, and butyric acid (4 carbons, in butter) has a sharp, pungent, acidic smell. (When the bioenergetic world talks about "free fatty acids" (FFAs) in the blood, it means these molecules floating around unbound, ready to be burned — or, if polyunsaturated, to cause trouble.)
In food and storage, three fatty acids are usually attached to a glycerol backbone, forming a triglyceride — the form fat is stored and eaten as. Digestion (the digestion page) snips the fatty acids off to use them. But the fatty acid itself is the molecule whose properties decide everything — so we now classify it.
The two axes — the model that fixes everyone's mental model
The most common mistake is to imagine fats on a single line from "short to long" or "good to bad." The reality is a two-dimensional grid, because fatty acids vary independently along two separate axes:
Axis 1 — Length (how many carbons in the tail)
You count the carbon atoms in the chain:
- Short-chain (SCFA): fewer than ~6 carbons — e.g. butyrate (4C, in butter and made by gut bacteria).
- Medium-chain (MCFA / "MCT"): ~6–12 carbons — e.g. caprylic (8C) and lauric (12C) acid, in coconut oil.
- Long-chain (LCFA): ~13–21 carbons — e.g. palmitic (16C, tallow), stearic (18C, cocoa butter), oleic (18C, olive oil). Most dietary fat is here.
- Very-long-chain (VLCFA): 22+ carbons.
Axis 2 — Saturation (how many double bonds in the tail)
"Saturation" asks: is the carbon chain "saturated" with hydrogen, or are some hydrogens missing? Carbon wants four bonds. If every carbon is holding its full quota of hydrogens, all the carbon-to-carbon bonds are single bonds and the chain is saturated. If hydrogens are missing, neighbouring carbons form a double bond (C=C) to compensate — and that double bond changes everything.
- Saturated (SFA) — zero double bonds. Every carbon full of hydrogen; only single bonds. The chain is straight and stiff — like a straight pencil — so the molecules pack together tightly and are solid at room temperature (butter, tallow, coconut oil). With no double bonds, they are chemically stable and resistant to oxidation.
- Monounsaturated (MUFA) — one double bond. Mono = one. That single double bond puts a rigid kink in the chain, so the molecules can't pack as tightly: liquid at room temperature but thickens in the fridge (olive oil, avocado, macadamia).
- Polyunsaturated (PUFA) — two or more double bonds. Multiple kinks make the molecule bend and curl, packing very poorly: liquid even when cold (fish oil, soybean, corn, sunflower oil). And — the crucial point for the whole second half of this page — those multiple double bonds make PUFA chemically reactive and prone to oxidation.
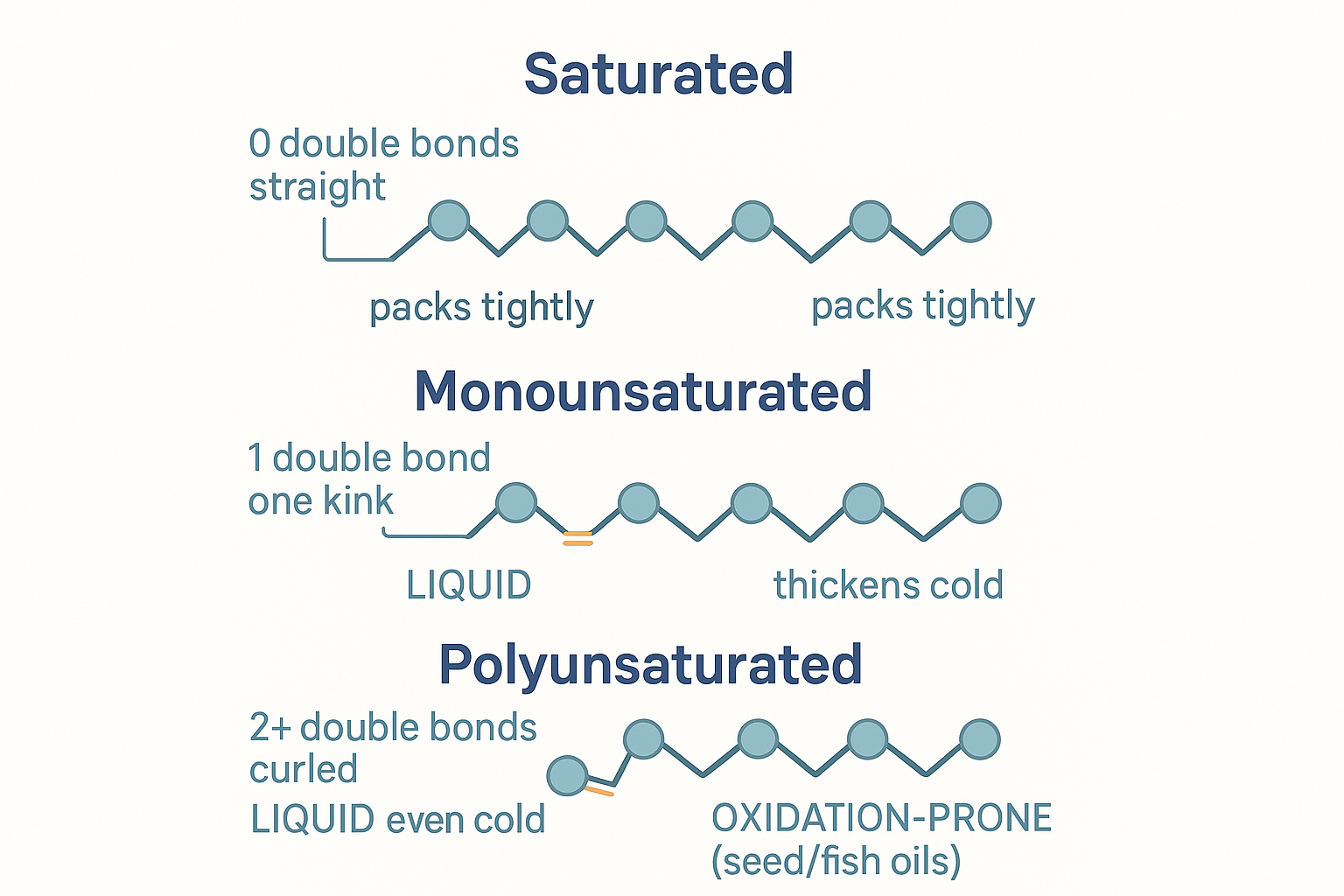 Saturation is shape: no double bonds = straight and stable; one = a kink; many = a curl that packs poorly and oxidises easily.
Saturation is shape: no double bonds = straight and stable; one = a kink; many = a curl that packs poorly and oxidises easily.
Putting the two axes together gives the grid that should replace the single-line mental model:
| Short / Medium chain | Long chain | |
|---|---|---|
| Saturated (0 double bonds) | Caprylic, lauric (MCT, coconut) | Palmitic (tallow), stearic (cocoa butter) |
| Monounsaturated (1 double bond) | rare in nature | Oleic (olive oil, macadamia) |
| Polyunsaturated (2+ double bonds) | rare in nature | Linoleic (omega-6, seed oils), EPA/DHA (omega-3, fish) |
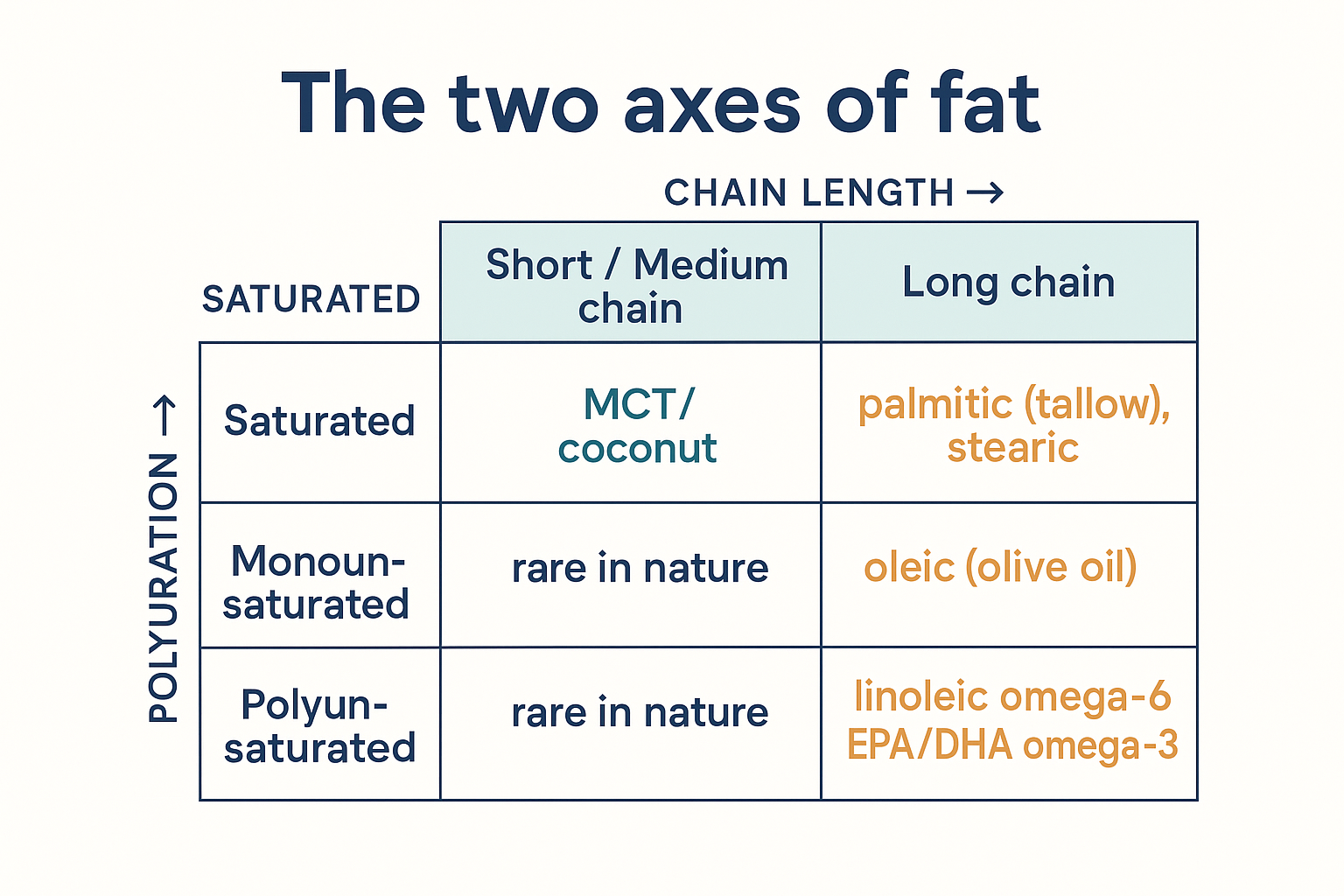 The mental model to adopt: fats live on a 2-D grid (length × saturation), not a single line.
The mental model to adopt: fats live on a 2-D grid (length × saturation), not a single line.
The omega number — where the first double bond sits
When you hear omega-3, omega-6, omega-9, it's simply describing the position of the first double bond, counting from the tail (methyl, "omega") end of the chain:
- Omega-3 — first double bond at the 3rd carbon from the tail (e.g. EPA/DHA in fish, ALA in flax).
- Omega-6 — first double bond at the 6th carbon (e.g. linoleic acid, the dominant fat in seed oils).
- Omega-9 — first at the 9th carbon (e.g. oleic acid, olive oil — note this one is a monounsaturated fat).
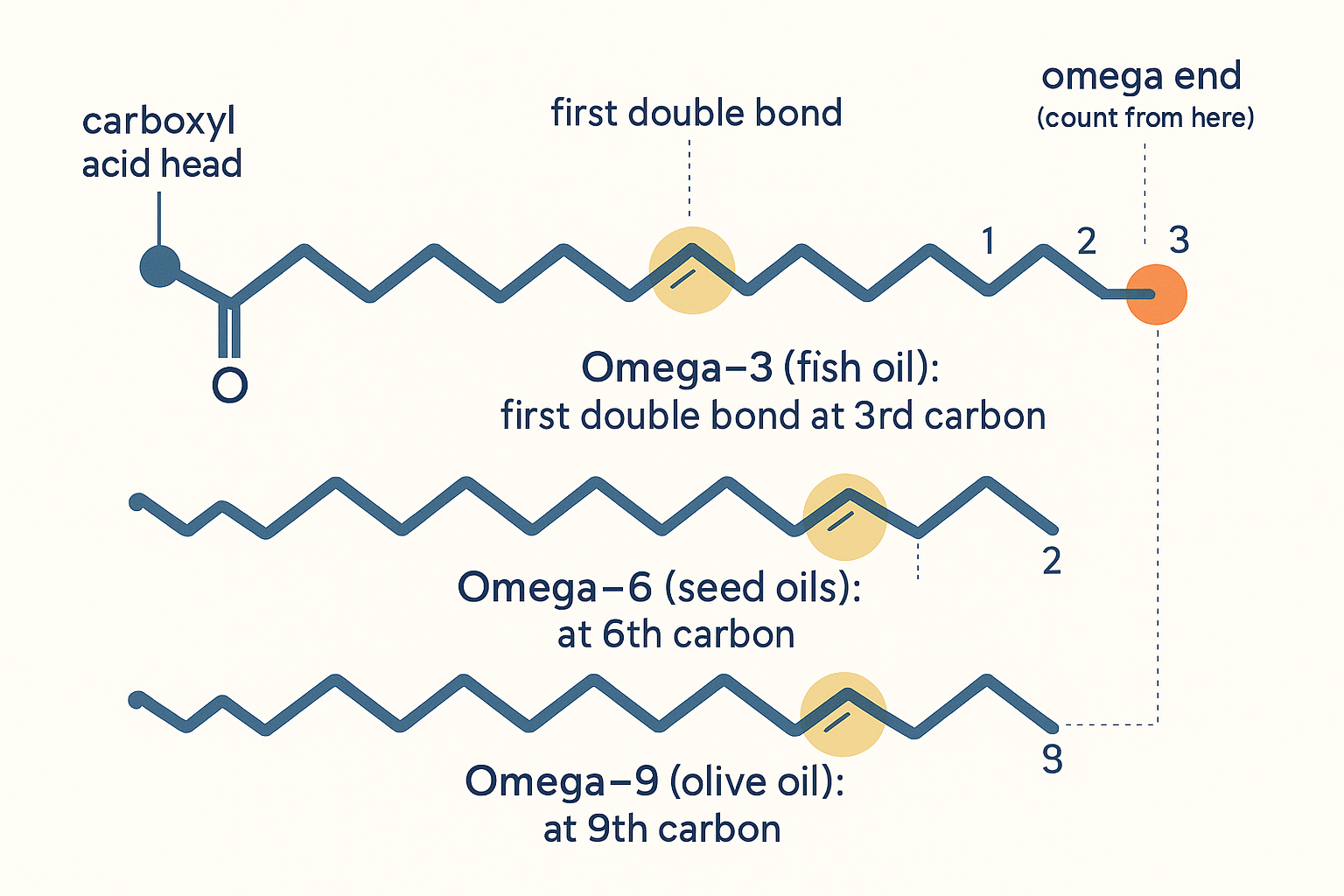 The omega number is just where the first double bond sits, counted from the tail end — a label for position, not a separate kind of fat.
The omega number is just where the first double bond sits, counted from the tail end — a label for position, not a separate kind of fat.
So "omega-X" is a sub-classification within unsaturated fats — a third descriptor, not a separate category. Omega-3 and omega-6 are both PUFAs; they differ in where the bonds start, which changes which signalling molecules they become (omega-6 tends toward inflammatory eicosanoids, omega-3 toward the pro-resolving ones from the inflammation page).
Trans fats — the unnatural outlier
One more category sits off this clean grid: trans fats. A natural unsaturated double bond is in the "cis" configuration — the chain kinks on the same side, giving that bend. Industrial partial hydrogenation (forcing hydrogen into liquid oils to make them solid and shelf-stable — margarine, old shortenings) flips some double bonds to the "trans" configuration, which is straight rather than kinked. The result is a molecule that's unsaturated chemically but packs and behaves like a saturated fat physically — and, critically, is foreign to human enzymes, which evolved for cis bonds. Trans fats are the one fat category with near-universal agreement that they are harmful (they worsen cholesterol profiles and inflammation and are now largely banned). They're the exception that proves the rule that shape and naturalness matter.
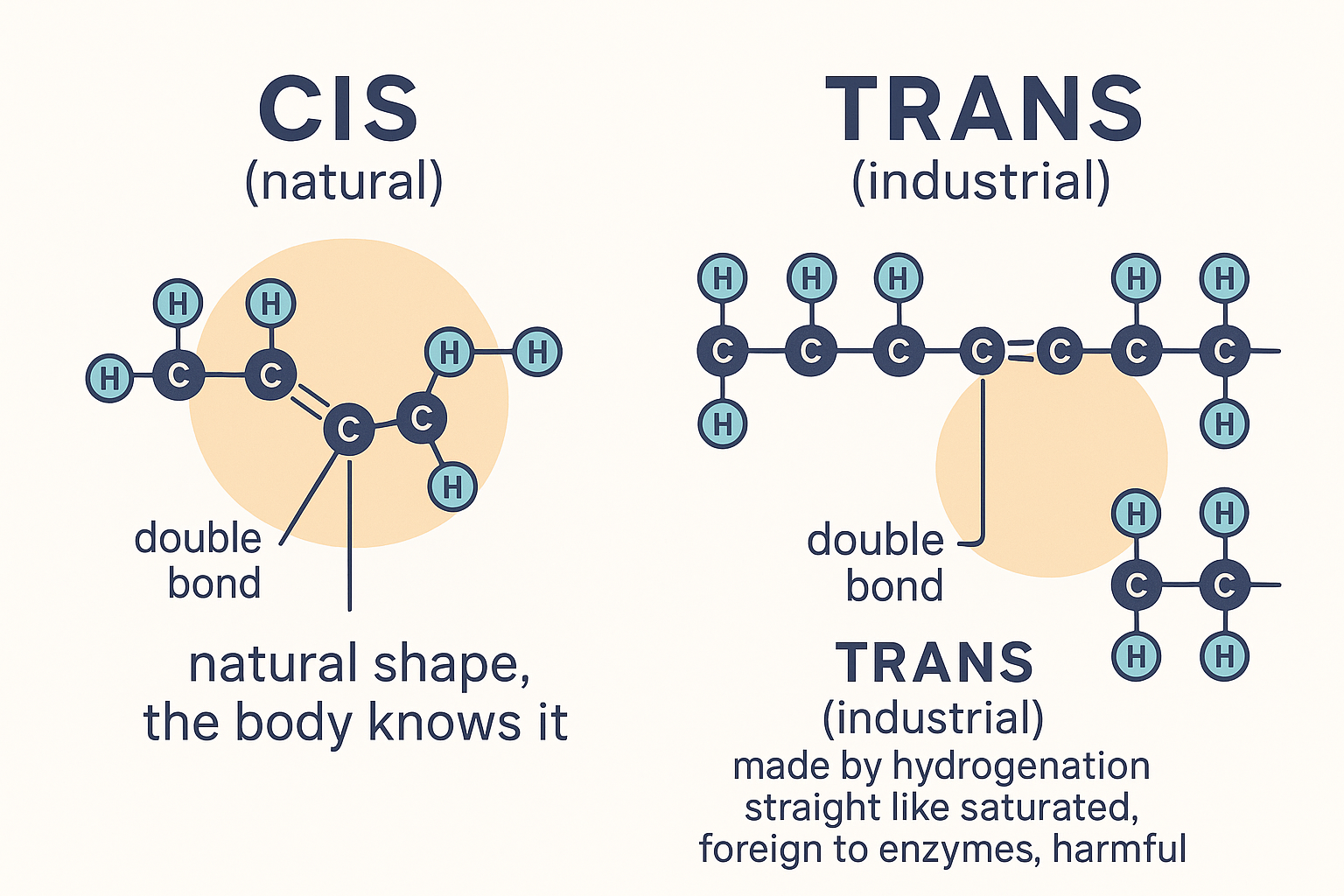 Same double bond, opposite shape: the natural cis bond bends the chain; the industrial trans bond leaves it straight — and foreign to your enzymes.
Same double bond, opposite shape: the natural cis bond bends the chain; the industrial trans bond leaves it straight — and foreign to your enzymes.
The fat types as second nature
Here is each type as a complete profile — chemistry, what happens in digestion, how it's metabolised, and its systemic/hormonal effect, with the why. This is the section to internalise.
Saturated fat (SFA) — e.g. butter, tallow, coconut, cocoa butter. Straight, stable, no double bonds. Digested and absorbed normally (long-chain SFAs via the lymphatic/chylomicron route, digestion page). Burned cleanly because it resists oxidation — it doesn't peroxidise on the way to the mitochondria. Structurally it makes membranes firmer. In the bioenergetic view it's the "safe" fat precisely because it has no reactive double bonds; mainstream concern is its effect on LDL cholesterol (real but more nuanced than "saturated fat = heart disease"). Why it's stable: no double bonds = no vulnerable points to oxidise.
Monounsaturated fat (MUFA) — e.g. olive oil, avocado, macadamia. One double bond — one vulnerable point, so moderately stable (much more than PUFA, less than SFA). Generally regarded as benign-to-beneficial across both mainstream and bioenergetic views. Oleic acid is also what your own body makes when it stores excess carbohydrate as fat. Why it's middling: one kink = one oxidation-prone site, a single risk rather than many.
Polyunsaturated fat (PUFA), omega-6 — e.g. linoleic acid, soybean/corn/sunflower oils. Two-plus double bonds → highly oxidation-prone (below). Becomes the substrate for inflammatory eicosanoids (arachidonic acid → COX/LOX → prostaglandins, the inflammation and aspirin pages). It is an essential fat (you need a little), but modern intake is enormous because seed oils saturate the food supply. The bioenergetic critique centres almost entirely here. Why it's the trouble-maker: many double bonds = many oxidation-prone sites + pro-inflammatory downstream products.
Polyunsaturated fat (PUFA), omega-3 — e.g. EPA/DHA in fish. Also polyunsaturated (even more double bonds — DHA has six), so also oxidation-prone and needing antioxidant protection. But its eicosanoid products are largely anti-inflammatory / pro-resolving (resolvins, inflammation page), so it's generally viewed positively — with the caveat (raised on the Vitamin E page) that it still raises your antioxidant requirement. A genuine tension: anti-inflammatory signalling vs peroxidation-proneness.
Medium-chain triglycerides (MCT) — e.g. caprylic/capric acid, coconut/MCT oil. Short enough (8–10C) to behave completely differently in metabolism: they bypass the normal fat-digestion machinery. From the digestion page, MCTs are absorbed directly into the portal vein (not packaged into chylomicrons and routed through lymph), going straight to the liver; and from the cellular energy page, they enter the mitochondria without the carnitine shuttle. So they're burned fast, are readily converted to ketones, and are saturated (stable). This is why MCT oil is the "fast, clean fat fuel." Why it's special: small size lets it skip both the slow lymphatic absorption route and the rate-limiting CPT1 gate.
Trans fat — e.g. partially hydrogenated oils (largely banned). Straight like saturated but foreign to your enzymes; interferes with normal fat metabolism and membrane function and is clearly pro-inflammatory and atherogenic. Avoid entirely. The one fat with no defenders.
flowchart TD
FA["A fatty acid"] --> L{How LONG?}
FA --> S{How SATURATED?}
L --> SC["Short (<6C): butyrate"]
L --> MC["Medium (6-12C): MCT/coconut<br/>→ fast, portal, ketones"]
L --> LC["Long (13-21C): most fat"]
S --> SFA["Saturated (0 C=C):<br/>straight · stable · solid"]
S --> MUFA["Mono (1 C=C):<br/>kinked · semi-stable"]
S --> PUFA["Poly (2+ C=C):<br/>curled · OXIDATION-PRONE"]
PUFA --> O6["omega-6: inflammatory"]
PUFA --> O3["omega-3: pro-resolving"]Part 2 — Why fat is energy, and how it's burned
Why fat is so caloric
Fat carries ~9 calories per gram, versus ~4 for carbohydrate and protein — more than double. The reason is molecular, and it's worth understanding because it also explains the peroxidation problem later.
A "calorie" is just stored chemical energy, and in food that energy lives almost entirely in carbon–hydrogen (C–H) bonds. Think of a C–H bond as a coiled spring: the electrons are held loosely and at high energy. "Burning" food means stripping hydrogens (and their electrons) off carbons and ultimately handing them to oxygen — an "electron hog" that holds electrons tightly at low energy. The drop from high-energy C–H to low-energy bonds-with-oxygen is the energy you harvest (as the NADH/FADH₂ → electron transport chain → ATP process from the cellular energy page).
Now compare the molecules:
- Carbohydrate (glucose) is already heavily oxygenated — the name carbo-hydrate hints at it; it's littered with oxygen-containing –OH groups. Many of its electrons are already in the low-energy, "partly-burned" state. It's like wood that's already half-charred.
- Fat is a long tail of almost pure carbon and hydrogen with virtually no oxygen — a dense, raw, completely unburned fuel rod. Gram for gram it packs far more harvestable C–H bonds.
So fat is calorie-dense because it's the most chemically reduced (oxygen-poor, hydrogen-rich) fuel — which also means burning it requires a lot of oxygen and, as we'll see, leaves more room for things to go wrong.
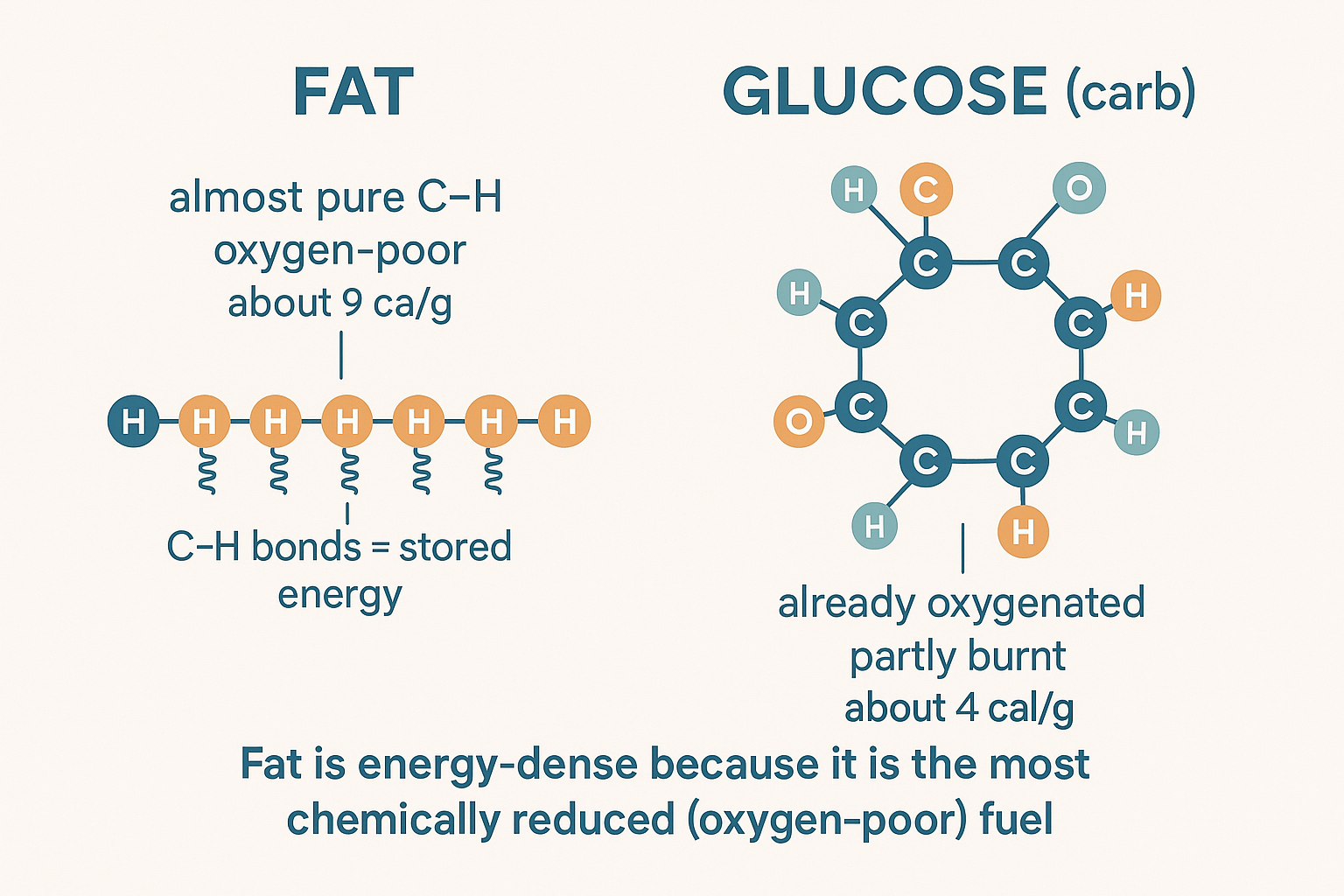 Why fat carries more than double the calories: it's almost pure C–H bonds with little oxygen, while glucose comes pre-oxygenated — already partly "burnt."
Why fat carries more than double the calories: it's almost pure C–H bonds with little oxygen, while glucose comes pre-oxygenated — already partly "burnt."
⚖️ Calibration. A common claim (in the AI explainer this page fact-checks) is that all fats have "the same calories." Close but not exact: they're all ~9 cal/g, but shorter chains (MCT) carry slightly fewer, and the tiny differences by saturation are negligible for practical purposes. "All roughly 9, MCT a bit less" is the accurate version.
How fat is actually burned: beta-oxidation in depth
The cellular energy page introduced this; here is the deeper version. To burn a long-chain fatty acid, the cell must (1) get it into the cell (CD36 transporter), (2) ferry it into the mitochondria via the carnitine shuttle (the CPT1 gate — the rate-limiting step, blocked by malonyl-CoA when sugar is plentiful), and then (3) run beta-oxidation.
Beta-oxidation is a repeating "chop" cycle: the enzyme machinery snips two carbons off the end of the fatty-acid chain at a time, and each chop yields:
- one acetyl-CoA (the two-carbon piece) → feeds the Krebs cycle,
- one FADH₂ and one NADH (electron carriers) → feed the electron transport chain directly.
A 16-carbon fat (palmitate) goes through this cycle seven times, producing 8 acetyl-CoA plus seven rounds of FADH₂/NADH — which is why fat yields so much ATP (~106 from one palmitate vs ~30 from glucose). The pathways then converge: fat-derived acetyl-CoA enters the same Krebs cycle and electron transport chain as glucose-derived acetyl-CoA.
flowchart LR
FAT["Long-chain fatty acid"] -->|CD36 into cell| CYT[In cytosol]
CYT -->|"carnitine shuttle (CPT1 gate)"| MAT[In mitochondrial matrix]
MAT --> BOX["β-oxidation:<br/>chop 2 carbons at a time"]
BOX -->|each chop| OUT["+1 acetyl-CoA<br/>+1 FADH2 +1 NADH"]
OUT --> KREBS[Krebs cycle + ETC<br/>= same as glucose]
MAT -.->|"when glucose scarce, liver makes"| KET[Ketones → brain fuel]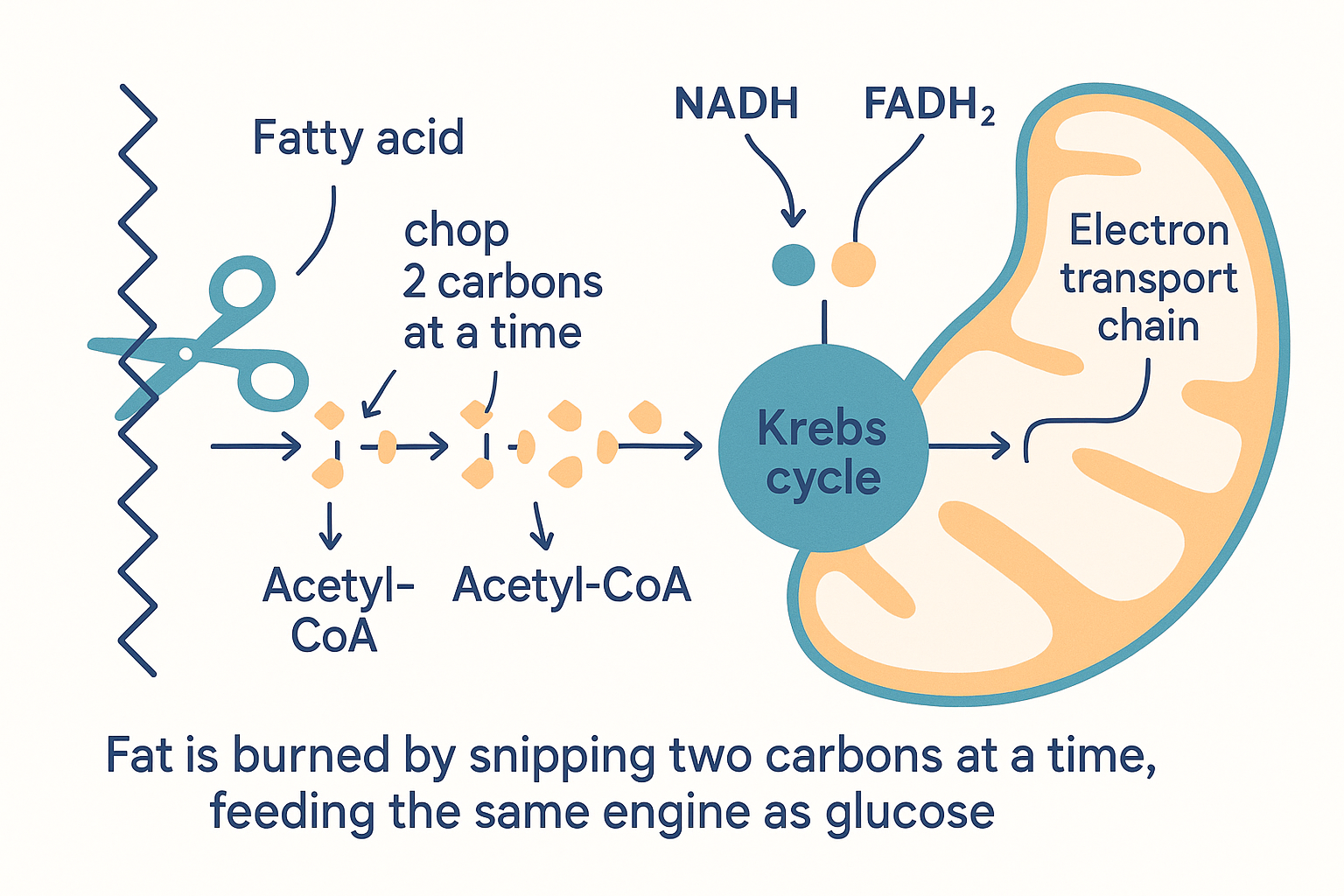 Burning fat means snipping the tail two carbons at a time — each snip yielding acetyl-CoA plus electron carriers that feed the same engine glucose uses.
Burning fat means snipping the tail two carbons at a time — each snip yielding acetyl-CoA plus electron carriers that feed the same engine glucose uses.
The FADH₂:NADH ratio — a real nuance, carefully stated. Different fuels deliver electrons to the chain in a different ratio of FADH₂ to NADH. Fatty-acid oxidation produces relatively more FADH₂ than glucose oxidation does. Because FADH₂ and NADH enter the electron transport chain at different points, this ratio affects how "backed up" the chain gets and how much reactive oxygen species (ROS) leak out — a higher FADH₂:NADH ratio (as from fat, especially saturated long-chain fat) tends to increase electron pressure and ROS production at one site.
⚖️ Calibration. The bioenergetic claim sometimes runs: "saturated fat's FADH₂:NADH ratio signals high metabolism." That's a stretch, and arguably backwards — a higher FADH₂:NADH ratio is associated with more reverse-electron-transport ROS, not a clean metabolic boost. The honest statement is: the ratio differs by fuel and influences ROS, which is a real and interesting effect — but it does not cleanly make any one fat "metabolically superior." Treat the F:N-ratio argument as a genuine but unsettled piece of biochemistry, not a slam-dunk.
Part 3 — The inflammatory dimension: why burning PUFA is playing with fire
This is the part the user most wanted demystified — the idea that broken fat creates "shards" that need vitamin E to clean up. Here is what that actually means, scientifically.
Lipid peroxidation: the chain reaction
Recall from Part 1 that a polyunsaturated fat has multiple double bonds, and that the carbons sitting between two double bonds (the bis-allylic positions) hold their hydrogens especially loosely. These are the weak points. When a stray reactive molecule (a free radical, or ROS — abundant wherever fuel is being burned) steals one of those loosely-held hydrogens, it turns that fat into a lipid radical, which grabs oxygen to become a lipid peroxyl radical, which then steals a hydrogen from the next fat molecule — turning it into a radical too.
That is the key: it's a self-propagating chain reaction. One initial hit can rip through a whole region of membrane, fat after fat, like a fuse burning along a row of molecules. This is lipid peroxidation, and it does two destructive things:
- It shreds the membrane the fats were forming — including mitochondrial membranes, damaging the very machinery burning the fuel.
- It produces toxic breakdown fragments — the "shards" — with names like 4-HNE (4-hydroxynonenal) and MDA (malondialdehyde). These are reactive aldehydes: small, mobile, and aggressive, they go on to damage proteins, enzymes, and DNA, and they are themselves inflammatory signals (they activate the NF-κB inflammatory pathway). So a peroxidised fat doesn't just damage its neighbourhood — it sends out reactive shrapnel that spreads damage and inflammation through the cell.
Why PUFA specifically: the number of bis-allylic weak points rises steeply with the number of double bonds. Saturated fat (no double bonds) has essentially zero such points and barely peroxidises; monounsaturated has few; polyunsaturated has many — and each additional double bond multiplies the vulnerability. This is the precise, mechanistic core of the entire "PUFA is uniquely dangerous" argument: more double bonds = more bis-allylic hydrogens = exponentially more peroxidation = more reactive aldehyde shards.
The antioxidant defence — where vitamin E comes in
The body isn't defenceless against this — it runs a chain-breaking antioxidant network, which is exactly the subject of the Vitamin E deep dive. The short version:
- Vitamin E sits inside the membrane among the fats and donates a hydrogen to the lipid peroxyl radical before it can attack the next fat — breaking the chain. It sacrifices itself, becoming a stable, harmless radical.
- Vitamin C and CoQ10/ubiquinol then regenerate spent vitamin E so it can work again — the cooperative antioxidant network.
- Glutathione (via glutathione peroxidase, needing selenium) mops up peroxides directly.
So the "shards require vitamin E and vitamin C" intuition is correct and mechanistic: the more PUFA you carry and burn, the more reactive peroxidation products you generate, and the more antioxidant capacity (vitamin E, C, CoQ10, glutathione) you consume defending against them. This is why the bioenergetic framework pairs lowering PUFA with keeping antioxidants high — two ends of the same problem.
flowchart LR
PUFA["PUFA in membrane<br/>(bis-allylic weak points)"] -->|ROS steals H| RAD[Lipid radical]
RAD -->|grabs oxygen| PEROX[Lipid peroxyl radical]
PEROX -->|"steals H from NEXT fat<br/>= CHAIN REACTION"| PUFA
PEROX --> SHARDS["Reactive aldehyde 'shards'<br/>4-HNE · MDA<br/>→ damage proteins/DNA<br/>→ activate inflammation"]
VITE["Vitamin E (in membrane)"] -->|"donates H, BREAKS chain"| PEROX
VITC["Vitamin C / CoQ10"] -->|regenerate| VITE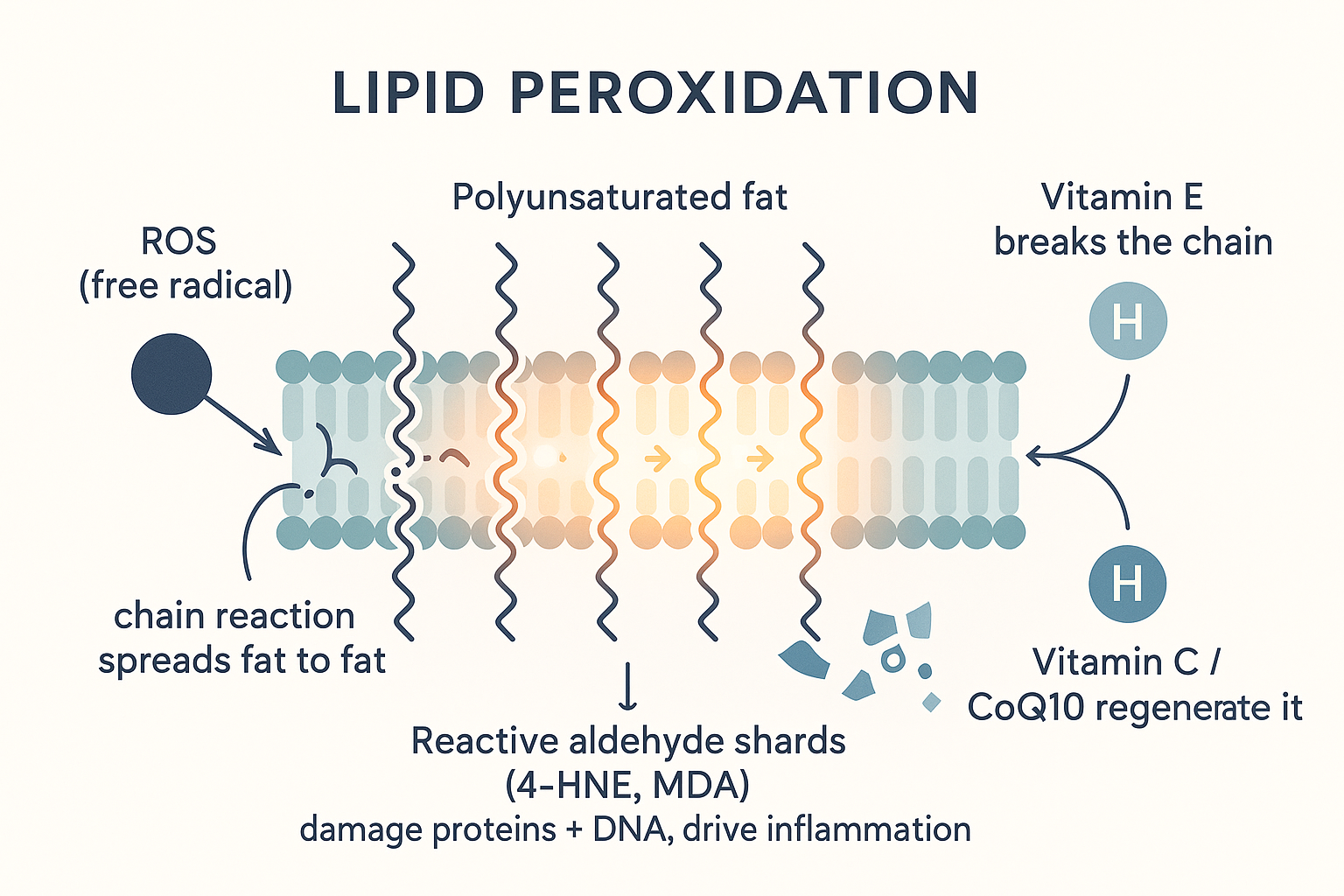 The "shards" made concrete: a single radical sets off a self-spreading fire through the membrane's PUFA, throwing off reactive aldehydes — until vitamin E steps in to break the chain.
The "shards" made concrete: a single radical sets off a self-spreading fire through the membrane's PUFA, throwing off reactive aldehydes — until vitamin E steps in to break the chain.
Part 4 — The Ray Peat cascade: why carbs may be the cleaner fuel
Now the framework the user asked to understand fully — not as a slogan but as an interdependent system. The naive intuition is "fat has more calories, so it's more energy, so it's better fuel." The bioenergetic counter-argument is that energy density (calories in the fuel) is not the same as metabolic efficiency (how cleanly the cell converts fuel to ATP without triggering a stress state) — and that carbohydrate, eaten in isolation, supports a higher metabolic state than fat eaten in isolation, through four linked mechanisms. Each is a real phenomenon; the interpretation of how much it matters is where the debate lives.
1. The CO₂ / oxygen-delivery angle
When you burn carbohydrate, you produce more carbon dioxide (CO₂) per unit of oxygen consumed than when you burn fat. This is just chemistry: carbs are more oxygenated to begin with, so their combustion yields a higher respiratory quotient (RQ ≈ 1.0 for carbs vs ≈ 0.7 for fat) — a well-established, non-controversial fact. The bioenergetic significance hangs on what CO₂ does: via the Bohr effect, higher CO₂ in the tissues makes haemoglobin release more oxygen into cells. So, somewhat counter-intuitively, burning carbs can improve tissue oxygenation. (Peat extended CO₂ into a "master regulator" with broader anti-stress, cell-stabilising roles — that broader claim is more his interpretation than settled consensus.)
2. The Randle cycle — fuels compete
From the cellular energy page: glucose and fat are wired as a see-saw (the Randle cycle). When free fatty acids rise in the blood, they actively suppress glucose oxidation — they inhibit the enzyme pyruvate dehydrogenase (PDH) that lets glucose-derived pyruvate into the mitochondria. So eating fat in isolation floods the blood with FFAs and forces the body into obligate fat-burning, shutting the door on glucose oxidation. The Peat framing: relying primarily on fat-burning is a "scarcity / hibernation" mode rather than a high-vitality state. (The Randle cycle is textbook; whether fat-burning is "worse" is the interpretive layer.)
3. Thyroid — the liver needs glucose to make active T3
From the liver and metabolism pages: the thyroid makes mostly inactive T4, which the liver converts to active T3 — and that conversion is supported by adequate liver glycogen (stored glucose). On a low-carbohydrate diet, liver glycogen runs low, and the body down-regulates T3 and produces more inactive reverse-T3 — slowing metabolic rate. This drop in T3 on low-carb/fasting is well-documented and real. The debate is whether it's a problem (a metabolic slowdown) or an adaptive efficiency — the keto world reads the same fact the opposite way.
4. The stress-hormone backstop
The brain is a glucose-greedy organ (cognition page) and cannot burn fatty acids directly. If you eat fat in isolation and blood sugar falls, the body must manufacture glucose (gluconeogenesis) to protect the brain — and it kick-starts that with stress hormones: cortisol and adrenaline. Cortisol breaks down tissue to make glucose; adrenaline mobilises fuel and can feel energising. Peat's framing: this is "stress-driven energy," not "thyroid-driven energy" — a body running on its emergency system. Carbohydrate, by contrast, feeds the brain directly and suppresses cortisol and adrenaline, signalling safety and permitting a high metabolic rate.
⚖️ Calibration — the honest caveats. This cascade is built from real phenomena (the RQ/CO₂ difference, the Randle cycle, the low-carb T3 drop, the gluconeogenic cortisol rise are all genuine). But two important corrections to the strong version: (a) the claim that the brain "cannot use fat" omits ketones — during sustained low-carb/fasting the liver makes ketones that fuel the brain substantially (50–70%), which blunts the glucose emergency and the stress-hormone reliance over time (keto-adaptation). The acute stress-hormone rise is real; the chronic picture is more adapted than the cascade implies. (b) Whether the lower-T3, fat-burning state is "worse" or simply "different/adaptive" is a genuine scientific disagreement, not a settled fact — the Peat framework treats high T3 and CO₂ as the goal, while the low-carb camp treats metabolic flexibility and fat-adaptation as the goal. This page presents the cascade so you understand the mechanisms; it does not declare a winner.
flowchart TD
FATISO["Eating FAT in isolation"] --> FFA[↑ Free fatty acids in blood]
FFA -->|Randle cycle: inhibits PDH| LOWGLU[↓ Glucose oxidation]
FATISO --> LOWGLY[↓ Liver glycogen]
LOWGLY -->|less substrate for conversion| LOWT3[↓ Active T3, ↑ reverse-T3]
FATISO --> LOWBS[Blood sugar falls]
LOWBS -->|brain needs glucose| GNG[Gluconeogenesis]
GNG --> STRESS[↑ Cortisol + adrenaline]
FATISO --> LOWCO2[↓ CO2 production<br/>less Bohr-effect O2 delivery]
LOWT3 --> STATE["Lower metabolic state<br/>(Peat's reading)"]
STRESS --> STATE
LOWGLU --> STATE
LOWCO2 --> STATE
LOWBS -.->|over time| KET["Ketones fuel brain<br/>(keto-adaptation blunts this)"]Putting it all together
- A fatty acid is a water-loving acid head (–COOH) on a water-fearing hydrocarbon tail — and it's classified on two independent axes: chain length (short/medium/long) and saturation (number of double bonds), with the omega number describing where the first double bond sits.
- Saturation determines shape, stability, and behaviour: saturated = straight, stable, solid; monounsaturated = one kink, semi-stable; polyunsaturated = many kinks, oxidation-prone; trans = unnatural straight, harmful; MCT = short enough to skip the slow absorption and carnitine gate and burn fast.
- Fat is calorie-dense (~9 cal/g) because it's the most chemically reduced fuel — almost pure C–H bonds with little oxygen, unlike already-oxygenated carbohydrate.
- It's burned by beta-oxidation (chop two carbons at a time → acetyl-CoA + FADH₂ + NADH → the same Krebs/ETC as glucose); the FADH₂:NADH ratio differs by fuel and affects ROS, but doesn't cleanly crown a "best" fat.
- Polyunsaturated fat is uniquely dangerous to carry and burn because its multiple double bonds create bis-allylic weak points that fuel a self-propagating lipid-peroxidation chain reaction, producing reactive aldehyde "shards" (4-HNE, MDA) that damage proteins/DNA and drive inflammation — which is exactly why it consumes vitamin E, C, and the antioxidant network.
- The bioenergetic case for carbs over fat rests on four real mechanisms (CO₂/Bohr oxygen delivery, the Randle cycle, glycogen-dependent T3 conversion, and the gluconeogenic cortisol/adrenaline backstop) — though ketones and keto-adaptation blunt the strong version, and whether the fat-burning state is "worse" or just "different" remains genuinely debated.
The unifying idea: "fat" is not one thing, and the arguments about it only make sense once you can see the molecule. Length and saturation set a fat's shape, stability, digestion, metabolism, and inflammatory potential; and the deepest divide in nutrition — fat vs carbohydrate as the body's preferred fuel — turns on whether you weight energy density or metabolic cleanliness. Understand the molecule and the cascade, and you can finally evaluate the claims yourself instead of taking sides.
Related compounds & foundations
Fats & fuels
- Coconut oil / MCTs — the fast, carnitine-skipping, ketogenic saturated fat.
- Fish oil — omega-3 PUFA: anti-inflammatory eicosanoids but still peroxidation-prone.
- Glucose — the competing fuel at the heart of the Randle cycle and the Peat cascade.
Antioxidant defence against peroxidation
- Vitamin E — the membrane chain-breaker (see the Vitamin E deep dive).
- Vitamin C, CoQ10 / ubiquinol — regenerate vitamin E.
- NAC / glutathione, selenium — the glutathione-peroxidase arm.
Metabolic levers
- Niacinamide, aspirin — anti-lipolytic tools that lower free fatty acids (the Randle-cycle and peroxidation angle).
- T3 — the active thyroid hormone the carbohydrate cascade protects.
Related foundations
- Cellular Energy — beta-oxidation, the carnitine shuttle, the Randle cycle, uncoupling.
- Inflammation — eicosanoids and the reactive products of lipid peroxidation.
- Systemic Metabolism — the thyroid and cortisol systems this page's cascade feeds into.
- Liver — glycogen, T4→T3 conversion, and fat processing.
- The Testosterone Kabbalah — the PUFA/estrogen/metabolism themes applied to hormones.